A Linnaeus NGTM Interactive Key to the Lithocolletinae of North-West Europe Aimed at Accelerating the Accumulation of Reliabl
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

1 Appendix 3. Thousand Islands National Park Taxonomy Report
Appendix 3. Thousand Islands National Park Taxonomy Report Class Order Family Genus Species Arachnida Araneae Agelenidae Agelenopsis Agelenopsis potteri Agelenopsis utahana Anyphaenidae Anyphaena Anyphaena celer Hibana Hibana gracilis Araneidae Araneus Araneus bicentenarius Larinioides Larinioides cornutus Larinioides patagiatus Clubionidae Clubiona Clubiona abboti Clubiona bishopi Clubiona canadensis Clubiona kastoni Clubiona obesa Clubiona pygmaea Elaver Elaver excepta Corinnidae Castianeira Castianeira cingulata Phrurolithus Phrurolithus festivus Dictynidae Emblyna Emblyna cruciata Emblyna sublata Eutichuridae Strotarchus Strotarchus piscatorius Gnaphosidae Herpyllus Herpyllus ecclesiasticus Zelotes Zelotes hentzi Linyphiidae Ceraticelus Ceraticelus atriceps 1 Collinsia Collinsia plumosa Erigone Erigone atra Hypselistes Hypselistes florens Microlinyphia Microlinyphia mandibulata Neriene Neriene radiata Soulgas Soulgas corticarius Spirembolus Lycosidae Pardosa Pardosa milvina Pardosa moesta Piratula Piratula canadensis Mimetidae Mimetus Mimetus notius Philodromidae Philodromus Philodromus peninsulanus Philodromus rufus vibrans Philodromus validus Philodromus vulgaris Thanatus Thanatus striatus Phrurolithidae Phrurotimpus Phrurotimpus borealis Pisauridae Dolomedes Dolomedes tenebrosus Dolomedes triton Pisaurina Pisaurina mira Salticidae Eris Eris militaris Hentzia Hentzia mitrata Naphrys Naphrys pulex Pelegrina Pelegrina proterva Tetragnathidae Tetragnatha 2 Tetragnatha caudata Tetragnatha shoshone Tetragnatha straminea Tetragnatha viridis -

Lepidoptera on the Introduced Robinia Pseudoacacia in Slovakia, Central Europe
Check List 8(4): 709–711, 2012 © 2012 Check List and Authors Chec List ISSN 1809-127X (available at www.checklist.org.br) Journal of species lists and distribution Lepidoptera on the introduced Robinia pseudoacacia in PECIES S OF ISTS L Slovakia, Central Europe Miroslav Kulfan E-mail: [email protected] Comenius University, Faculty of Natural Sciences, Department of Ecology, Mlynská dolina B-1, SK-84215 Bratislava, Slovakia. Abstract: Robinia pseudoacacia A current checklist of Lepidoptera that utilize as a hostplant in Slovakia (Central Europe) faunalis provided. community. The inventory Two monophagous is based on species, a bibliographic the leaf reviewminers andMacrosaccus new unreported robiniella data and from Parectopa southwest robiniella Slovakia., and Thethe polyphagouslist includes 35pest Lepidoptera Hyphantria species cunea belonging to 10 families. Most species are polyphagous and belong to Euro-Siberian have subsequently been introduced to Slovakia. Introduction E. The area is a polygon enclosed by the towns of Bratislava, Robinia pseudoacacia a widespread species in its native habitat in southeastern North America. It was L.introduced (black locust, to orEurope false acacia),in 1601 is Komárno, Veľký Krtíš and Myjava. Ten plots were located in the southern part of the study area. Most were located in theThe remnant trophic ofgroups the original of the floodplain Lepidoptera forests larvae that found were (Chapman 1935). The first mention of planting the species distributed along the Danube and Morava rivers. (Keresztesiin Slovakia dates 1965). from Today, 1750, itwhen is widespread black locust wasthroughout planted (1986). The zoogeographical distribution of the species western,around the central, fortress eastern in Komárno and southern in southern Europe, Slovakia where followswere defined the arrangement following the give system by Reiprichof Brown (2001). -

Phyllonorycter Issikii
EUROPEAN AND MEDITERRANEAN PLANT PROTECTION ORGANIZATION ЕВРОПЕЙСКАЯ И СРЕДИЗЕМНОМОРСКАЯ ОРГАНИЗАЦИЯ ПО КАРАНТИНУ И ЗАЩИТЕ РАСТЕНИЙ ORGANIZATION EUROPEENNE ET MEDITERRANEENNE POUR LA PROTECTION DES PLANTES Data Sheets on Forest Pests Phyllonorycter issikii IDENTITY Name: Phyllonorycter issikii (Kumata) Synonym: Lithocolletis issikii Kumata Taxonomic position: Insecta: Lepidoptera, Gracillariidae Common name: Lime leaf miner (English); Липовая минирующая моль-пестрянка (Russian). Bayer computer code: PRYCIS HOSTS Larvae of P. issikii make folded mines in the lower side of leaves of Tilia spp (preferred host). The native hosts in the Far East are, T cordata, T. amurensis, T. mandshurica, T. maximowicziana and other Tilia, but also Betula platyphylla (Ermolaev, 1977; Kozlov, 1991; Orlinskii et al., 1991b). In the West of Russia, Tilia cordata is the preferred host (and also where it is introduced in the East). P.issikii has not specifically been recorded on Tilia platyphyllas or the hybrid T europea, most widely oplanted in western Europe. GEOGRAPHICAL DISTRIBUTION EPPO region: Lithuania (recently introduced), Russia (South of the Far East; South and centre of the European part – introduced: cities of Voronezh, Samara, Ufa, Moscow and their vicinities), Ukraine (introduced). Asia: Korea, Russia (South of the Far East), Japan (Kumata et al., 1983; Kozlov, 1991; Orlinskii et al., 1991). EU: Absent. BIOLOGY The caterpillars of the first generation of P. issikii pupate in the second half of June; moths fly from the end of June till the middle of July. The second generation develops from the end of July till the end of August; moths fly from the middle of August till the beginning of September. Folded mines are located on the lower side of leaves. -

Checkered Beetles Moths (Lepidoptera: Gracillariidae) – Hazardous Phytophags of Arboreal and Shrubby Plants of Botanical Gardens and Plantings of Kiev M
UDC 632.634.791.937 (477.75) © 2017 Checkered beetles moths (Lepidoptera: Gracillariidae) – hazardous phytophags of arboreal and shrubby plants of botanical gardens and plantings of Kiev M. Lisovyi, O. Sylchuk Natsional University of Life and Environmental Sciences of Ukraine, Heroev Oborony str., 13, Kyiv, 03041, Ukraine P. Chumak, V. Kovalchuk, Botanichny Garden of Acad. O. Fomina The purpose. To carry out probes on revealing and specification of species composition of checkered moths (Lepidoptera: Gracillariidae) in conditions of botanical gardens and plantings of Kiev. Methods. Standard methods of faunistic research in entomology, population ecology, and protection of plants. Results. It is determined that 24 kinds of checkered moths are eating 54 kinds of plants which are widely used for gardening in Kiev. For the first time the following kinds are revealed: Phyllonorycter issikii, Phyllonorycter platani, and Phyllonorycter emberizaepennella. At calculation of Palii-Kovnatski indexes they specified that in city plantings the dominant phytophags are Cameraria ohridella (94,11%), Phyllonorycter populifoliella (86,37%) and Gracillaria syringella (59,14%). They consider that in formation of the secondary areal of invasion kinds of checkered moths the great value has an areal of spread of the host-plant. Environmental analysis is carried out of checkered moths of family Gracillariidae which is spread in cities of the Europe and which are absent in fauna of cities of Ukraine. That has important theoretical and practical value for ecology, entomology and protection of plants against hazardous checkered moths. Conclusions. All the probed kinds of checkered moths by their trophic specialization may be distributed into polyphages (6 kinds), oligophages (14 kinds) and monophages (3 kinds). -

A New Leaf-Mining Moth from New Zealand, Sabulopteryx Botanica Sp
A peer-reviewed open-access journal ZooKeys 865: 39–65A new (2019) leaf-mining moth from New Zealand, Sabulopteryx botanica sp. nov. 39 doi: 10.3897/zookeys.865.34265 MONOGRAPH http://zookeys.pensoft.net Launched to accelerate biodiversity research A new leaf-mining moth from New Zealand, Sabulopteryx botanica sp. nov. (Lepidoptera, Gracillariidae, Gracillariinae), feeding on the rare endemic shrub Teucrium parvifolium (Lamiaceae), with a revised checklist of New Zealand Gracillariidae Robert J.B. Hoare1, Brian H. Patrick2, Thomas R. Buckley1,3 1 New Zealand Arthropod Collection (NZAC), Manaaki Whenua–Landcare Research, Private Bag 92170, Auc- kland, New Zealand 2 Wildlands Consultants Ltd, PO Box 9276, Tower Junction, Christchurch 8149, New Ze- aland 3 School of Biological Sciences, The University of Auckland, Private Bag 92019, Auckland, New Zealand Corresponding author: Robert J.B. Hoare ([email protected]) Academic editor: E. van Nieukerken | Received 4 March 2019 | Accepted 3 May 2019 | Published 22 Jul 2019 http://zoobank.org/C1E51F7F-B5DF-4808-9C80-73A10D5746CD Citation: Hoare RJB, Patrick BH, Buckley TR (2019) A new leaf-mining moth from New Zealand, Sabulopteryx botanica sp. nov. (Lepidoptera, Gracillariidae, Gracillariinae), feeding on the rare endemic shrub Teucrium parvifolium (Lamiaceae), with a revised checklist of New Zealand Gracillariidae. ZooKeys 965: 39–65. https://doi.org/10.3897/ zookeys.865.34265 Abstract Sabulopteryx botanica Hoare & Patrick, sp. nov. (Lepidoptera, Gracillariidae, Gracillariinae) is described as a new species from New Zealand. It is regarded as endemic, and represents the first record of its genus from the southern hemisphere. Though diverging in some morphological features from previously de- scribed species, it is placed in genus Sabulopteryx Triberti, based on wing venation, abdominal characters, male and female genitalia and hostplant choice; this placement is supported by phylogenetic analysis based on the COI mitochondrial gene. -

Hymenoptera: Eulophidae) 321-356 ©Entomofauna Ansfelden/Austria; Download Unter
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Entomofauna Jahr/Year: 2007 Band/Volume: 0028 Autor(en)/Author(s): Yefremova Zoya A., Ebrahimi Ebrahim, Yegorenkova Ekaterina Artikel/Article: The Subfamilies Eulophinae, Entedoninae and Tetrastichinae in Iran, with description of new species (Hymenoptera: Eulophidae) 321-356 ©Entomofauna Ansfelden/Austria; download unter www.biologiezentrum.at Entomofauna ZEITSCHRIFT FÜR ENTOMOLOGIE Band 28, Heft 25: 321-356 ISSN 0250-4413 Ansfelden, 30. November 2007 The Subfamilies Eulophinae, Entedoninae and Tetrastichinae in Iran, with description of new species (Hymenoptera: Eulophidae) Zoya YEFREMOVA, Ebrahim EBRAHIMI & Ekaterina YEGORENKOVA Abstract This paper reflects the current degree of research of Eulophidae and their hosts in Iran. A list of the species from Iran belonging to the subfamilies Eulophinae, Entedoninae and Tetrastichinae is presented. In the present work 47 species from 22 genera are recorded from Iran. Two species (Cirrospilus scapus sp. nov. and Aprostocetus persicus sp. nov.) are described as new. A list of 45 host-parasitoid associations in Iran and keys to Iranian species of three genera (Cirrospilus, Diglyphus and Aprostocetus) are included. Zusammenfassung Dieser Artikel zeigt den derzeitigen Untersuchungsstand an eulophiden Wespen und ihrer Wirte im Iran. Eine Liste der für den Iran festgestellten Arten der Unterfamilien Eu- lophinae, Entedoninae und Tetrastichinae wird präsentiert. Mit vorliegender Arbeit werden 47 Arten in 22 Gattungen aus dem Iran nachgewiesen. Zwei neue Arten (Cirrospilus sca- pus sp. nov. und Aprostocetus persicus sp. nov.) werden beschrieben. Eine Liste von 45 Wirts- und Parasitoid-Beziehungen im Iran und ein Schlüssel für 3 Gattungen (Cirro- spilus, Diglyphus und Aprostocetus) sind in der Arbeit enthalten. -
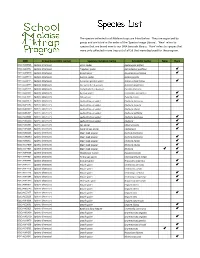
Species List
The species collected in all Malaise traps are listed below. They are organized by group and are listed in the order of the 'Species Image Library'. ‘New’ refers to species that are brand new to our DNA barcode library. 'Rare' refers to species that were only collected in one trap out of all 59 that were deployed for the program. -

Redalyc.Interactions Among Host Plants, Lepidoptera Leaf Miners And
SHILAP Revista de Lepidopterología ISSN: 0300-5267 [email protected] Sociedad Hispano-Luso-Americana de Lepidopterología España Yefremova, Z. A.; Kravchenko, V. D. Interactions among host plants, Lepidoptera leaf miners and their parasitoids in the forest- steppe zone of Russia (Insecta: Lepidoptera, Hymenoptera) SHILAP Revista de Lepidopterología, vol. 43, núm. 170, junio, 2015, pp. 271-280 Sociedad Hispano-Luso-Americana de Lepidopterología Madrid, España Available in: http://www.redalyc.org/articulo.oa?id=45541421012 How to cite Complete issue Scientific Information System More information about this article Network of Scientific Journals from Latin America, the Caribbean, Spain and Portugal Journal's homepage in redalyc.org Non-profit academic project, developed under the open access initiative 271-280 Interactions among host 3/6/15 10:45 Página 271 SHILAP Revta. lepid., 43 (170), junio 2015: 271-280 eISSN: 2340-4078 ISSN: 0300-5267 Interactions among host plants, Lepidoptera leaf miners and their parasitoids in the forest-steppe zone of Russia (Insecta: Lepidoptera, Hymenoptera) Z. A. Yefremova & V. D. Kravchenko Abstract The article reports on the quantitative description of the food web structure of the community consisting of 65 species of Lepidoptera leaf miners reared from 34 plant species, as well as 107 species of parasitoid eulophid wasps (Hymenoptera: Eulophidae). The study was conducted in the forest-steppe zone of the Middle Volga in Russia over 13 years (2000-2012). Leaf miners have been found to be highly host plant-specific. Most of them are associated with only one or two plant species and therefore the number of links between trophic levels is 73, which is close to the total number of Lepidoptera species (linkage density is 1.12). -

Parasitoids of Phyllonorycter Platani (Staudinger) (Lepidoptera, Gracillariidae) in Serbia
www.ccsenet.org/jps Journal of Plant Studies Vol. 1, No. 1; March 2012 Parasitoids of Phyllonorycter platani (Staudinger) (Lepidoptera, Gracillariidae) in Serbia Čedomir Marković (Corresponding author) Faculty of Forestry, Belgrade University Kneza Višeslava 1, Belgrade 11030, Serbia Tel: 381-113-553-122 E-mail: [email protected] Aleksandar Stojanović Natural History Museum Njegoševa 51, Belgrade 11000, Serbia Tel: 381-113-442-147 E-mail: [email protected] Received: September 8, 2011 Accepted: October 23, 2011 Published: March 1, 2012 doi:10.5539/jps.v1n1p79 URL: http://dx.doi.org/10.5539/jps.v1n1p79 Abstract In the study of parasitoids of the plane leaf miner Phyllonorycter platani during 2004, 18 polyphagous parasitoid species were recorded in four tree rows and four parks in Serbia (3 species of the family Ichneumonidae and 15 species of the family Eulophidae). Among the recorded parasitoids, the most significant were the species Minotetrastichus platanellus, Pediobius saulius and M. frontalis, followed by the species Pnigalio agraules, P. pectinicornis, Sympiesis gordius and Neochrysocharis formosa. The other parasitoid species were rare and few. The parasitism rates in the study samples of Ph. platani ranged from 15.0 to 37.8 %. Keywords: Parasitism, Parasitoids, Phyllonorycter platani, Platanus acerifolia 1. Introduction Thanks to its high ornamental value, fast growth, resistance to air pollution and dust, European plane (Platanus acerifolia (Ait.) Willd.) is frequent in parks and tree rows in Serbia. Unfortunately, its trees are often severely infested by the leaf miner Phyllonorycter platani. For that reason, the planes loose much of their ornamental value and their premature defoliations are frequent (Marković et al., 2006). -

Annual Report 2011
Annual Report 2011 © RMCA www.africamuseum.be Foreword 2 Foreword The Royal Museum for Central Africa (RMCA) pub- and culture exhibition was extended, while RMCA lishes a beautiful and richly illustrated annual collection pieces were admired in more than 20 report in book form every two years. In intervening major exhibitions held in different parts of the years – such as 2011 – we publish a digital edition globe. Nearly 30,000 children attended our edu- that is available on our website, and for which a cational workshops or school activities, while our hard copy can be produced on demand. Despite colla boration with African communities became its size, the report is not exhaustive. Rather, it more streamlined. We felt a pang of regret at the seeks to provide the most varied overview pos- departure of ‘our’ elephants in 2011. After grac- sible of our many museum-related, educational, ing our museum’s entrance for three years, the scientific, and other activities on the national and 9 pachyderms that formed the work created by international scene. The long governmental crisis South African artist Andries Botha, You can buy my of 2011 notwithstanding, RMCA was highly pro- heart and my soul, left Tervuren Park for good. ductive and remains one of the most important Africa-focused research institutions, particularly 2011 was also a fruitful year in terms of scientific for Central Africa. research. To highlight the multidisciplinary nature that is the strength of our institution, we organ- As with the previous year, the renovation was one ized ‘Science Days’ for the first time. -

Pheromone Trap Catch of Harmful Microlepidoptera Species in Connection with the Chemical Air Pollutants
International Journal of Research in Environmental Science (IJRES) Volume 2, Issue 1, 2016, PP 1-10 ISSN 2454-9444 (Online) http://dx.doi.org/10.20431/2454-9444.0201001 www.arcjournals.org Pheromone Trap Catch of Harmful Microlepidoptera Species in Connection with the Chemical Air Pollutants L. Nowinszky1, J. Puskás1, G. Barczikay2 1University of West Hungary Savaria University centre, Szombathely, Károlyi Gáspár Square 4 2County Borsod-Abaúj-Zemplén Agricultural Office of Plant Protection and Soil Conservation Directorate, Bodrogkisfalud, Vasút Street. 22 [email protected], [email protected] Abstract: In this study, seven species of Microlepidoptera pest pheromone trap collection presents the results of the everyday function of the chemical air pollutants (SO2, NO, NO2, NOx, CO, PM10, O3). Between 2004 and 2013 Csalomon type pheromone traps were operating in Bodrogkisfalud (48°10’N; 21°21E; Borsod-Abaúj- Zemplén County, Hungary, Europe). The data were processed of following species: Spotted Tentiform Leafminer (Phyllonorycter blancardella Fabricius, 1781), Hawthorn Red Midged Moth (Phyllonorycter corylifoliella Hübner, 1796), Peach Twig Borer (Anarsia lineatella Zeller, 1839), European Vine Moth (Lobesia botrana Denis et Schiffermüller, 1775), Plum Fruit Moth (Grapholita funebrana Treitschke, 1846), Oriental Fruit Moth (Grapholita molesta Busck, 1916) and Codling Moth (Cydia pomonella Linnaeus, 1758). The relation is linear, logarithmic and polynomial functions can be characterized. Keywords: Microlepidoptera, pests, pheromone traps, air pollution 1. INTRODUCTION Since the last century, air pollution has become a major environmental problem, mostly over large cities and industrial areas (Cassiani et al. 2013). It is natural that the air pollutant chemicals influence the life phenomena of insects, such as flight activity as well. -

The Cassidinae Beetles of Longnan County (Jiangxi, China): Overview and Community Composition
Biodiversity Data Journal 7: e39053 doi: 10.3897/BDJ.7.e39053 Research Article The Cassidinae beetles of Longnan County (Jiangxi, China): overview and community composition Peng Liu‡, Chengqing Liao‡‡, Jiasheng Xu , Charles L. Staines§, Xiaohua Dai ‡,| ‡ Leafminer Group, School of Life Sciences, Gannan Normal University, Ganzhou, China § Smithsonian Environmental Research Center, Edgewater, United States of America | National Navel-Orange Engineering Research Center, Ganzhou, China Corresponding author: Xiaohua Dai ([email protected]) Academic editor: Flávia Rodrigues Fernandes Received: 13 Aug 2019 | Accepted: 16 Oct 2019 | Published: 18 Oct 2019 Citation: Liu P, Liao C, Xu J, Staines CL, Dai X (2019) The Cassidinae beetles of Longnan County (Jiangxi, China): overview and community composition. Biodiversity Data Journal 7: e39053. https://doi.org/10.3897/BDJ.7.e39053 Abstract There are few reports on the community composition and diversity pattern of the Cassidinae species of China. Compared to the neighbouring provinces of Guangdong, Fujian and Zhejiang, the Cassidinae richness in Jiangxi Province is under-reported. Longnan City, a biodiversity hotspot in Jiangxi Province, was chosen to obtain the first overview of the Cassidinae beetles. The sample coverage curves for the three sample sites reached an asymptote which indicated sampling was sufficient for data analysis. A total of eight tribes, 16 genera, 59 species and 1590 individuals of Cassidinae beetles were collected. Most belonged to the tribe Hispini (1121 individuals; 70.5%), followed by the tribe Cassidini (161 individuals; 10.13%) and the tribe Oncocephalini (159 individuals; 10.0%). The remainder (149 individuals) belonged to five tribes (Gonophorini, Basiprionotini, Callispini, Notosacanthini and Aspidimorphini). The tribes Notosacanthini, Aspidimorphini and Oncocephalini were newly recorded for Jiangxi Province.