Differential Effects of Temperature Variability on the Transmission of a Marine Parasite
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-
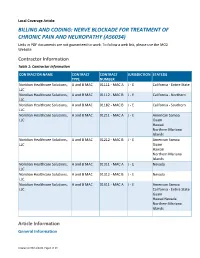
BILLING and CODING: NERVE BLOCKADE for TREATMENT of CHRONIC PAIN and NEUROPATHY (A56034) Links in PDF Documents Are Not Guaranteed to Work
Local Coverage Article: BILLING AND CODING: NERVE BLOCKADE FOR TREATMENT OF CHRONIC PAIN AND NEUROPATHY (A56034) Links in PDF documents are not guaranteed to work. To follow a web link, please use the MCD Website. Contractor Information Table 1: Contractor Information CONTRACTOR NAME CONTRACT CONTRACT JURISDICTION STATE(S) TYPE NUMBER Noridian Healthcare Solutions, A and B MAC 01111 - MAC A J - E California - Entire State LLC Noridian Healthcare Solutions, A and B MAC 01112 - MAC B J - E California - Northern LLC Noridian Healthcare Solutions, A and B MAC 01182 - MAC B J - E California - Southern LLC Noridian Healthcare Solutions, A and B MAC 01211 - MAC A J - E American Samoa LLC Guam Hawaii Northern Mariana Islands Noridian Healthcare Solutions, A and B MAC 01212 - MAC B J - E American Samoa LLC Guam Hawaii Northern Mariana Islands Noridian Healthcare Solutions, A and B MAC 01311 - MAC A J - E Nevada LLC Noridian Healthcare Solutions, A and B MAC 01312 - MAC B J - E Nevada LLC Noridian Healthcare Solutions, A and B MAC 01911 - MAC A J - E American Samoa LLC California - Entire State Guam Hawaii Nevada Northern Mariana Islands Article Information General Information Created on 05/11/2021. Page 1 of 23 Article ID A56034 Article Title Billing and Coding: Nerve Blockade for Treatment of Chronic Pain and Neuropathy Article Type Billing and Coding AMA CPT / ADA CDT / AHA NUBC Copyright Statement CPT codes, descriptions and other data only are copyright 2020 American Medical Association. All Rights Reserved. Applicable FARS/HHSARS apply. Fee schedules, relative value units, conversion factors and/or related components are not assigned by the AMA, are not part of CPT, and the AMA is not recommending their use. -

Journeys to Byzantium? Roman Senators Between Rome and Constantinople
Journeys to Byzantium? Roman Senators Between Rome and Constantinople Master’s Thesis Presented in Partial Fulfillment of the Requirements for the Degree Master of Arts in the Graduate School of The Ohio State University By Michael Anthony Carrozzo, B.A Graduate Program in History The Ohio State University 2010 Thesis Committee: Kristina Sessa, Advisor Timothy Gregory Anthony Kaldellis Copyright by Michael Anthony Carrozzo 2010 Abstract For over a thousand years, the members of the Roman senatorial aristocracy played a pivotal role in the political and social life of the Roman state. Despite being eclipsed by the power of the emperors in the first century BC, the men who made up this order continued to act as the keepers of Roman civilization for the next four hundred years, maintaining their traditions even beyond the disappearance of an emperor in the West. Despite their longevity, the members of the senatorial aristocracy faced an existential crisis following the Ostrogothic conquest of the Italian peninsula, when the forces of the Byzantine emperor Justinian I invaded their homeland to contest its ownership. Considering the role they played in the later Roman Empire, the disappearance of the Roman senatorial aristocracy following this conflict is a seminal event in the history of Italy and Western Europe, as well as Late Antiquity. Two explanations have been offered to explain the subsequent disappearance of the Roman senatorial aristocracy. The first involves a series of migrations, beginning before the Gothic War, from Italy to Constantinople, in which members of this body abandoned their homes and settled in the eastern capital. -

The Edictum Theoderici: a Study of a Roman Legal Document from Ostrogothic Italy
The Edictum Theoderici: A Study of a Roman Legal Document from Ostrogothic Italy By Sean D.W. Lafferty A thesis submitted in conformity with the requirements for the degree of Doctor of Philosophy Department of History University of Toronto © Copyright by Sean D.W. Lafferty 2010 The Edictum Theoderici: A Study of a Roman Legal Document from Ostrogothic Italy Sean D.W. Lafferty Doctor of Philosophy Department of History University of Toronto 2010 Abstract This is a study of a Roman legal document of unknown date and debated origin conventionally known as the Edictum Theoderici (ET). Comprised of 154 edicta, or provisions, in addition to a prologue and epilogue, the ET is a significant but largely overlooked document for understanding the institutions of Roman law, legal administration and society in the West from the fourth to early sixth century. The purpose is to situate the text within its proper historical and legal context, to understand better the processes involved in the creation of new law in the post-Roman world, as well as to appreciate how the various social, political and cultural changes associated with the end of the classical world and the beginning of the Middle Ages manifested themselves in the domain of Roman law. It is argued here that the ET was produced by a group of unknown Roman jurisprudents working under the instructions of the Ostrogothic king Theoderic the Great (493-526), and was intended as a guide for settling disputes between the Roman and Ostrogothic inhabitants of Italy. A study of its contents in relation to earlier Roman law and legal custom preserved in imperial decrees and juristic commentaries offers a revealing glimpse into how, and to what extent, Roman law survived and evolved in Italy following the decline and eventual collapse of imperial authority in the region. -

The Fifth Century, the Decemvirate, and the Quaestorship
THE FIFTH CENTURY, THE DECEMVIRATE, AND THE QUAESTORSHIP Ralph Covino (University of Tennessee at Chattanooga). There has always been some question as to the beginnings of the regularized quaestorship. As long ago as 1936, Latte pointed out that there seems to be no Roman tradition as to the quaestors’ origins.1 As officials, they barely appear at all in the early Livian narrative. The other accounts which we possess, such as those of Tacitus and Ioannes Lydus, are unhelpful at best. While scholars such as Lintott and others have sought to weave together the disparate fragments of information that exist so as to construct a plausible timeline of events surrounding the first appearance of regular, elected quaestors in the middle of the fifth-century, it becomes apparent that there is a dimension that has been omitted. Magisterial offices, even ones without the power to command and to compel, imperium, do not appear fully-formed overnight. This paper seeks to uncover more of the process by which the regular quaestors emerged so as to determine how they actually came to be instituted and regularized rather than purely rely on that which we are told happened from the sources.2 Tacitus records that quaestors were instituted under the monarchs in a notoriously problematic passage: The quaestorship itself was instituted while the kings still reigned, as shown by the renewal of the curiate law by L. Brutus; and the power of selection remained with the consuls, until this office, with the rest, passed into the bestowal of the people. The first election, sixty-three years after the expulsion of the Tarquins, was that of Valerius Potitus and Aemilius Mamercus, as finance officials attached to the army in the field. -
MORSE HALL CB N Floor # Fitness for a Particular Use
X170 150A 196 LOADING DN DOCK 180 170 150 UP DN X142 190F 190C 142 140 UP 190A 194 193 192 181 190B 170B 190E 170A 165 160 UP 190D 197 141 195 191 190 198 Building/Zone Western Washington University (WWU) makes these documents available on an "as is" basis. All warranties and representations of any kind with regard to said 0 8 16 documents are disclaimed, including the implied warranties of merchantability and TRUE MORSE HALL CB N Floor # fitness for a particular use. WWU does not warrant the documents against FT NORTH FLOOR 1 deficiencies of any kind. Date Revised: 12/13/2016 Last Sync With FAMIS: 09/22/2015 1 270B 270C 270A 275 270 270D 296 280 281 DN UP 290D 260 254 282 265 258 256 252 250 290N 245 283 284 290C 290W 244 290A 202 295 229 295A 230B 230A 201 210A 220A 290S 243 294 242 293 292 200 210 220 230 241 292A UP 291 200A 240 290E X290E Building/Zone Western Washington University (WWU) makes these documents available on an "as is" basis. All warranties and representations of any kind with regard to said 0 8 16 documents are disclaimed, including the implied warranties of merchantability and TRUE MORSE HALL CB N Floor # fitness for a particular use. WWU does not warrant the documents against FT NORTH FLOOR 2 deficiencies of any kind. Date Revised: 12/13/2016 Last Sync With FAMIS: 09/22/2015 2 386 385 387B 387A 383 390G 387 DN UP 396 382 380 370 360 350 388 390N 381 345 389 390C 390W 344 390S 395 310B 329 395A 310C 310A 330A 343 394 393 342 392 310 330 330B 341 392A UP 391 340 DN 390E ROOF BELOW BASE FLOOR PLAN Building Western Washington University (WWU) makes these documents available on an "as is" basis. -

3.2 Precipitation Or Dry-Wet Reconstructions
Climate change in China during the past 2000 years: An overview Ge Quansheng , Zheng Jingyun Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China Email: [email protected] Outline 1 Introduction 2 Historical Documents as Proxy 3 Reconstructions and Analyses 4 Summary and Prospects 1. Introduction: E Asia2K Climate System Socio-economic System •typical East Asian •dense population and rapid monsoon climate economic development • significant seasonal and • be susceptible to global inter-annual and inter- warming and extreme decadal variability climate events Climate change study in the past 2ka in East Asian is both beneficial and advantageous. • various types of natural proxy • Plenty of historical documents Fig. Active regional working groups under as proxy the past 2ka theme (PAGES 2009) 2. Historical Documents as Proxy Type Period Amount Chinese classical 1,531 kinds, 137 BC~1470 AD documents 32,251 volumes More than 8,000 1471~1911 (The Ming Local gazettes books (部), 110, and Qing Dynasty) 000 volumes Memos to the About 120,000 1736~1911 emperor pieces Archives of the 1912~1949 20,000 volumes Republic of China More than 200 Private diaries 1550~ books (部) Chinese classical documents AD 833, North China plain: Extreme drought event was occurred, crops were shriveling, no yields, people were in hungry…. Fig. Example for Ancient Chinese writings Local gazettes The 28th year of the Daoguang reign (1848 AD), the 6th (lunar) month, strong wind and heavy rain, the Yangtze River overflowed; the 7th month, strong wind Fig. Gazettes of Yangzhou Prefecture and thunder storm, field published in 1874 AD and houses submerged. -

Hospitalitas
Department of Economics Working Paper Series Hospitalitas Andrew T. Young Working Paper No. 15-41 This paper can be found at the College of Business and Economics Working Paper Series homepage: http://be.wvu.edu/phd_economics/working-papers.htm Hospitalitas† Andrew T. Young College of Business and Economics West Virginia University Morgantown, WV 26506-6025 ph: 304 293 4526 em: [email protected] Latest Version: October 2015 Abstract: Good government requires a constitution that demarcates what political agents can and cannot do, and such a constitution must be self-enforcing. The medieval West was characterized by the estates system, where the political power of monarchs was roughly balanced by that of a landed and militarized nobility. This rough balance of power contributed to a Western tradition of limited government and constitutional bargaining. I argue that this balance has important roots in the fifth and sixth century barbarian settlements that occurred within the frontiers of the declining Western Roman Empire. These settlements provided barbarians with allotments consisting of lands or claims to taxes due from those lands. These allotments aligned the incentives of barbarian warriors and Roman landowners; they also realigned (or newly aligned) the incentives of barbarian warriors and leadership elite as their roving confederacies became stationary kingdoms. Barbarian military forces became decentralized and the warriors became political powerful shareholders of the realm. JEL Codes: H10, P16, P48, N40, N43, Keywords: constitutional political economy, polycentric sovereignty, shareholder states, collective action problems, governance institutions, state emergence † I thank the participants at the 2015 Western Economic Association International meetings in Honolulu, Hawaii for valuable comments and suggestions. -

Chapter Thirteen New Covenant and Other Christianities, to Ca. 185 CE
Chapter Thirteen New Covenant and Other Christianities, to ca. 185 CE The disastrous revolt of 66-70 marks a sharp turning point in the history of both Judaism and Christianity. The militancy of the first Christiani did not survive the disaster (that an oracle ordered “the Christians” to flee from Jerusalem to Pella just before the war of 66-70 is a much later invention).1 Two very different directions taken in the generation after the destruction of the Jerusalem temple led to the dramatic growth of two distinct religions: New Covenant Christianity and rabbinic Judaism. Although they had a common heritage these two religions differed widely not only from each other but also from the religion of Judaea during the Second Temple period. Rabbinic Judaism devoted itself fervently to the torah, in the belief that Adonai had punished Judaea because its people had been too lax in following his law. New Covenant Christians, contrarily, believed that God had ordained the destruction of the temple to make way for the parousia of Jesus the Christ and the End of Time. That religiosity and superstitious fanaticism were the cause of the disaster did not occur to either camp. A small minority of Judaeans, however, followed a third course in the aftermath of 70: these people gave up entirely on Adonai, his covenants, and the temporal world, and formulated - on the authority, they claimed, of Jesus the Christ - a mythical explanation of reality that scholars call “Gnosticism.” Although we have more information about each of the three new movements than about traditional Judaism in the Greek-speaking Diaspora, it must be added that for a very long time after 70 CE the majority of Judaeans did not adhere to any one of the three. -

From the Holy Land to the Cloister: the Decline of Female Ascetic Pilgrimages in the Early Medieval West (C
From the Holy Land to the Cloister: The Decline of Female Ascetic Pilgrimages in the Early Medieval West (c. 350-615) Manon Claire Williams Department of History University of Colorado at Boulder Primary Thesis Advisor Dr. Scott Bruce, Department of History Honors Council Representative Dr. Fred Anderson, Department of History Outside Reader Dr. Andrew Cain, Department of Classics Defense: April 2nd 2015 Abstract This paper will focus on the mobility of ascetic women from late antiquity through to the early Middle Ages with a particular emphasis on the practice of pilgrimage. As seen in multiple primary source documents, religious women from the West were journeying to the Holy Land and beyond from the fourth through to the early fifth centuries. This practice, however, is mentioned remarkably less in accounts of religious women north of the Alps in the late fifth century onwards. Evidence of women undertaking pilgrimages to the Holy Land is sparse while their male counterparts continued to make such journeys. Although the monastic rules that cloistered women certainly did have an impact on the movement of religious women in the early medieval West as other scholars have argued, there are a variety of external and non-religious factors that also contributed to the decline of female ascetic pilgrimages. This thesis will aim to give a more complete answer to this decline by focusing on the functions that religious women played in their communities, which effectively bound them to those same communities. Due to a lack of source material, the main focus of the early medieval period will be in Gaul, which will provide a case study for understanding the roles that religious women played in society within the context of early Merovingian politics and economy. -
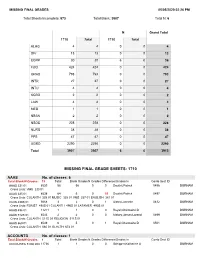
Missing Final Grade Sheets: 1710
MISSING FINAL GRADES 05/05/2020 02:26 PM Total Sheets Incomplete: 573 Total Blank: 3907 Total N: 6 N Grand Total 1710 Total 1710 Total ALHG 4 4 0 0 4 DIV 13 13 0 0 13 EGRP 30 30 6 6 36 FUQ 424 424 0 0 424 GRAD 793 793 0 0 793 INTR 27 27 0 0 27 INTU 4 4 0 0 4 KGRD 2 2 0 0 2 LAW 4 4 0 0 4 MED 1 1 0 0 1 NBSN 2 2 0 0 2 NSOE 228 228 0 0 228 NURS 38 38 0 0 38 PPS 47 47 0 0 47 UGRD 2290 2290 0 0 2290 Total 3907 3907 6 6 3913 MISSING FINAL GRADE SHEETS: 1710 AAAS No. of classes: 6 Total Blank/N Grades: 73 Total Blank Grades N Grades Difference/Grades In Comb Sect ID AAAS 331 01 8535 58 58 0 0 Douthit,Patrick 0496 DURHAM Cross Lists: VMS 230 01 AAAS 335 01 8538 64 5 0 59 Douthit,Patrick 0497 DURHAM Cross Lists: CULANTH 335 01 MUSIC 335 01 VMS 337 01 ENGLISH 381 01 AAAS 490S 01 5668 3 2 0 1 Aidoo,Lamonte 0432 DURHAM Cross Lists: ROMST 490S 01 CULANTH 490S 01 LATAMER 490S 01 AAAS 496 01 11271 1 1 0 0 Royal,Charmaine D DURHAM AAAS 512S 01 8543 2 2 0 0 Matory,James Lorand 0499 DURHAM Cross Lists: CULANTH 511S 01 RELIGION 511S 01 AAAS 660 01 8549 6 5 0 1 Royal,Charmaine D 0501 DURHAM Cross Lists: CULANTH 660 01 GLHLTH 672 01 ACCOUNTG No. -

The Enduring Legacy of the Iatrosophist Gessius Edward Watts
The Enduring Legacy of the Iatrosophist Gessius Edward Watts HE IATROSOPHIST GESSIUS offers historians of later Roman intellectual history a peculiar but remarkably Tdurable historical profile. A native of Petra, Gessius made a name for himself by practicing and teaching medicine in Alexandria in the late fifth and early sixth centuries.1 Both Christian and pagan sources speak highly of his skills as a physician, skills which earned him a rare collection of honors from the Roman state.2 Gessius, however, plays an incidental role in the history of the larger Roman world. He never en- joyed the fame of Galen, the political influence of Oribasius,3 the diplomatic prominence of Uranius,4 or even the literary 1 For his origin in Petra, Dam. Isid. fr.128 Athanassiadi. The Suda places his floruit in the reign of Zeno (Γ 207), a date confirmed by (and probably derived from) Damascius’ portrait. The recollection of Gessius by Ste- phanus of Athens suggests that Gessius may have been teaching into the 530s (L. G. Westerink, Stephanus of Athens: Commentary on Hippocrates’ Aphorisms I [CMG XI.1.3.1 (Berlin 1985)] 20). On Gessius’ dates see also the earlier comments of O. Temkin, “Geschichte des Hippokratismus im ausgehenden Altertum,” Kyklos 4 (1932) 1–80, at 73–74. 2 His teaching earned for him χρημάτων μεγάλων ἐγένετο κύριος καὶ Ῥωμαϊκῶν ἔτυχεν ἀξιωμάτων οὐ τῶν τυχόντων (Dam. Isid. fr.128). 3 See B. Baldwin, “The Career of Oribasius,” Acta Classica 18 (1975) 85– 97, and “Beyond the House Call: Doctors in Early Byzantine History and Politics,” DOP 38 (1984) 15–19, at 17. -

Download Free at ISBN 978‑1‑909646‑72‑8 (PDF Edition) DOI: 10.14296/917.9781909646728
Ravenna its role in earlier medieval change and exchange Ravenna its role in earlier medieval change and exchange Edited by Judith Herrin and Jinty Nelson LONDON INSTITUTE OF HISTORICAL RESEARCH Published by UNIVERSITY OF LONDON SCHOOL OF ADVANCED STUDY INSTITUTE OF HISTORICAL RESEARCH Senate House, Malet Street, London WC1E 7HU First published in print in 2016 (ISBN 978‑1‑909646‑14‑8) This book is published under a Creative Commons Attribution‑ NonCommercial‑NoDerivatives 4.0 International (CC BY‑ NCND 4.0) license. More information regarding CC licenses is available at https://creativecommons.org/licenses/ Available to download free at http://www.humanities‑digital‑library.org ISBN 978‑1‑909646‑72‑8 (PDF edition) DOI: 10.14296/917.9781909646728 iv Contents Acknowledgements vii List of contributors ix List of illustrations xiii Abbreviations xvii Introduction 1 Judith Herrin and Jinty Nelson 1. A tale of two cities: Rome and Ravenna under Gothic rule 15 Peter Heather 2. Episcopal commemoration in late fifth‑century Ravenna 39 Deborah M. Deliyannis 3. Production, promotion and reception: the visual culture of Ravenna between late antiquity and the middle ages 53 Maria Cristina Carile 4. Ravenna in the sixth century: the archaeology of change 87 Carola Jäggi 5. The circulation of marble in the Adriatic Sea at the time of Justinian 111 Yuri A. Marano 6. Social instability and economic decline of the Ostrogothic community in the aftermath of the imperial victory: the papyri evidence 133 Salvatore Cosentino 7. A striking evolution: the mint of Ravenna during the early middle ages 151 Vivien Prigent 8. Roman law in Ravenna 163 Simon Corcoran 9.