Re-Evaluation of the Placentia Bay-Grand Banks Area to Identify Ecologically and Biologically Significant Areas
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Broadcasting Notice of Consultation CRTC 2012-212
Broadcasting Notice of Consultation CRTC 2012-212 PDF version Ottawa, 10 April 2012 Notice of hearing 7 June 2012 Gatineau, Quebec Deadline for submission of interventions/comments/answers: 10 May 2012 [Submit an intervention/comment/answer or view related documents] The Commission will hold a hearing commencing on 7 June 2012 at 2 p.m. at the Commission Headquarters, 1 Promenade du Portage, Gatineau, Quebec. The Commission intends to consider, subject to interventions, the following applications without the appearance of the parties: Applicant/licensee and locality 1. MOTV Média Inc. Across Canada Application 2012-0170-7 2. MOTV Média Inc. Across Canada Application 2012-0171-4 3. Rogers Broadcasting Limited Across Canada Application 2012-0173-0 4. 3924181 Canada Inc. Across Canada Application 2012-0197-0 5. Larry C. Osmond Grand Bank, Newfoundland and Labrador Application 2011-0969-5 6. Colba.Net Telecom Inc. Fredericton, Moncton, Saint John, Allardville, Big Cove, Blue Mountain Settlement, Bouctouche, Brown’s Flat, Burtts Corner, Cap Lumière, Davis Mill, Caron Brook, Centre-Acadie, Centre Napan, Clair, Harvey, Highway 505/St-Édouard, Jacquet River, Keating’s Corner, Lac Baker, Ludford Subdivision, McAdam, Morrisdale, Musquash Subdivision, Nasonworth, Noonan, Patterson/Hoyt, Petitcodiac, Richibucto, Ruchibucto Village, 2 Rogersville, St-André-de-Shediac, Ste-Anne-de-Kent, St-Antoine, St-Ignace, St-Joseph-de-Madawaska, Ste-Marie-de-Kent, Salmon Beach, Tracy/Fredericton Junction, Welsford, Willow Grove and their surrounding areas in New Brunswick; St. John’s, Deer Lake, Pasadena and their surrounding areas in Newfoundland and Labrador; Dartmouth, Halifax, Bedford, Sackville and their surrounding areas in Nova Scotia Application 2012-0174-8 7. -

Replacement Class Screening Report
REPLACEMENT CLASS SCREENING WORKS ON OVER-WINTERING SITES FOR OYSTER AQUACULTURE REPLACEMENT CLASS SCREENING WORKS ON OVER-WINTERING SITES FOR OYSTER AQUACULTURE TRANSPORT CANADA ENVIRONMENTAL AFFAIRS ATLANTIC REGION Heritage Court 95 Foundry Street Moncton, NB E1C 8K6 ii TABLE OF CONTENTS 4.2. ANALYSIS AND PREDICTION OF SIGNIFICANCE OF RESIDUAL ENVIRONMENTAL EFFECTS ....................................... 14 1. INTRODUCTION ......................................................................1 5. ISSUES SCOPING AND VALUED ENVIRONMENTAL 1.1. CLASS SCREENING AND THE CANADIAN COMPONENT SELECTION ................................................................. 15 ENVIRONMENTAL ASSESSMENT ACT (CEAA) ...................................3 5.1. ANALYSIS OF ENVIRONMENTAL EFFECTS ON 1.2. RATIONALE FOR REPLACEMENT CLASS SELECTED VECS ................................................................................. 15 SCREENING (RCS).................................................................................4 5.2. MARINE HABITAT (MARINE WATERS AND MARINE 1.3. CONSULTATION................................................................5 SEDIMENTS)........................................................................................ 15 1.4 CANADIAN ENVIRONMENTAL ASSESSMENT 5.2.1. WILDLIFE/MIGRATORY BIRDS................................... 16 REGISTRY (THE REGISTRY)....................................................................5 5.2.2. SPECIES AT RISK.......................................................... 16 2. PROJECTS SUBJECT TO CLASS -

The Hitch-Hiker Is Intended to Provide Information Which Beginning Adult Readers Can Read and Understand
CONTENTS: Foreword Acknowledgements Chapter 1: The Southwestern Corner Chapter 2: The Great Northern Peninsula Chapter 3: Labrador Chapter 4: Deer Lake to Bishop's Falls Chapter 5: Botwood to Twillingate Chapter 6: Glenwood to Gambo Chapter 7: Glovertown to Bonavista Chapter 8: The South Coast Chapter 9: Goobies to Cape St. Mary's to Whitbourne Chapter 10: Trinity-Conception Chapter 11: St. John's and the Eastern Avalon FOREWORD This book was written to give students a closer look at Newfoundland and Labrador. Learning about our own part of the earth can help us get a better understanding of the world at large. Much of the information now available about our province is aimed at young readers and people with at least a high school education. The Hitch-Hiker is intended to provide information which beginning adult readers can read and understand. This work has a special feature we hope readers will appreciate and enjoy. Many of the places written about in this book are seen through the eyes of an adult learner and other fictional characters. These characters were created to help add a touch of reality to the printed page. We hope the characters and the things they learn and talk about also give the reader a better understanding of our province. Above all, we hope this book challenges your curiosity and encourages you to search for more information about our land. Don McDonald Director of Programs and Services Newfoundland and Labrador Literacy Development Council ACKNOWLEDGMENTS I wish to thank the many people who so kindly and eagerly helped me during the production of this book. -
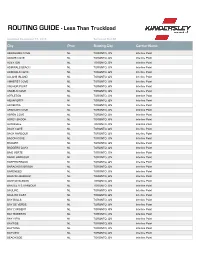
ROUTING GUIDE - Less Than Truckload
ROUTING GUIDE - Less Than Truckload Updated December 17, 2019 Serviced Out Of City Prov Routing City Carrier Name ABRAHAMS COVE NL TORONTO, ON Interline Point ADAMS COVE NL TORONTO, ON Interline Point ADEYTON NL TORONTO, ON Interline Point ADMIRALS BEACH NL TORONTO, ON Interline Point ADMIRALS COVE NL TORONTO, ON Interline Point ALLANS ISLAND NL TORONTO, ON Interline Point AMHERST COVE NL TORONTO, ON Interline Point ANCHOR POINT NL TORONTO, ON Interline Point ANGELS COVE NL TORONTO, ON Interline Point APPLETON NL TORONTO, ON Interline Point AQUAFORTE NL TORONTO, ON Interline Point ARGENTIA NL TORONTO, ON Interline Point ARNOLDS COVE NL TORONTO, ON Interline Point ASPEN COVE NL TORONTO, ON Interline Point ASPEY BROOK NL TORONTO, ON Interline Point AVONDALE NL TORONTO, ON Interline Point BACK COVE NL TORONTO, ON Interline Point BACK HARBOUR NL TORONTO, ON Interline Point BACON COVE NL TORONTO, ON Interline Point BADGER NL TORONTO, ON Interline Point BADGERS QUAY NL TORONTO, ON Interline Point BAIE VERTE NL TORONTO, ON Interline Point BAINE HARBOUR NL TORONTO, ON Interline Point BAKERS BROOK NL TORONTO, ON Interline Point BARACHOIS BROOK NL TORONTO, ON Interline Point BARENEED NL TORONTO, ON Interline Point BARR'D HARBOUR NL TORONTO, ON Interline Point BARR'D ISLANDS NL TORONTO, ON Interline Point BARTLETTS HARBOUR NL TORONTO, ON Interline Point BAULINE NL TORONTO, ON Interline Point BAULINE EAST NL TORONTO, ON Interline Point BAY BULLS NL TORONTO, ON Interline Point BAY DE VERDE NL TORONTO, ON Interline Point BAY L'ARGENT NL TORONTO, ON -

A Brief History of the Random Region of Trinity Bay
A Brief History of the Random Region of Trinity Bay A Presentation to the Wessex Society St. John's, Nfld. Leslie J. Dean April, 1994 Revised June 16, 1997 HISTORY OF THE RANDOM REGION OF TRINITY BAY INTRODUCTION The region encompassed by the Northwest side of Trinity Bay bounded by Southwest Arm (of Random), Northwest Arm (of Random), Smith's Sound and Random Island has, over the years, been generally referred to as "Random". However, "Random" is now generally interpreted locally as that portion of the region encompassing Southwest Arm and Northwest Arm. Effective settlement of the region as a whole occurred largely during the 1857 - 1884 period. A number of settlements at the outer fringes of the region including Rider's Harbour at the eastern extremity of Random Island and "Harts Easse" at the entrance to Random Sound were settled much earlier. Indeed, these two settlements together with Ireland's Eye near the eastern end of Random Island, were locations of British migratory fishing activity in Trinity Bay throughout the l600s and 1700s. "Harts Easse" was the old English name for Heart's Ease Beach. One of the earliest references to the name "Random" is found on the 1689 Thornton's map of Newfoundland which shows Southwest Arm as River Random. It is possible that the region's name can be traced to the "random" course which early vessels took when entering the region from the outer reaches of Trinity Bay. In all likelihood, however, the name is derived from the word random, one meaning of which is "choppy" or "turbulence", which appropriately describes 2 sea state conditions usually encountered at the region's outer headlands of West Random Head and East Random Head. -

S Storm-Petrels on Baccalieu Island, Newfoundland
POPULATION STUDY HEBREEDING RANGE OF LEACH'S Storm-Petrel( Oceanodroma leu- The world'slargest known corhoa) in the Northwest Atlan- tic is centered in eastern Newfound- land. Owing to the species'nocturnal nestingcolony of and burrow-nestinghabits, the breed- ing populations and distributions of storm-petrels, first documented in Leach's Storm-Petrels Newfoundlandby Petersand Burleigh (1951), have historicallybeen grossly underestimatedand are not presently on BaccalieuIsland,, well known. Storm-petrel coloniesin Newfoundland are vastly larger than all others in the Northwest Atlantic Newfoundland (Huntington 1963; Montevecchi, in prep.), but few quantitative data on colony populationshave been gener- ated. The colony of Leach's Storm-Pe- trels on Baccalieu Island, Newfound- B. O. Sklepkovych and IV. A. Montevecchi land, hasgone largely unrecognized as an important nestingsite for the spe- des (Brown et al. 1975; Nettleship 1980). In the present paper, we esti- mate the breeding population of storm-petrelsbased on measurements of burrow densitiesin all nestinghab- itats and on quantitative estimatesof the island'ssurface areas. Our popu- lation figure, which is compared with thoseof other coloniesthroughout the world, yields new perspectivesof the energy consumption of these birds in the marine ecosystemof the North- west Atlantic. Study Area BaccalieuIsland (-6.3 x I kilome- ter, 48'07'N, 54ø12'W) is situatedoff the northeastern tip of Newfound- land's Avalon Peninsula (Figure 1). Leach'sStorm-Petrels nest in a variety of habitats over the entire island, but principallyoccupy grassy-hummock- ed slopes,boreal forest and Empetrum heaths. Smaller, less defined habitats suchas herb (grassyareas with more than 60 percent fern or tall herb cover), grassy meadow, and heath Leach• Storm-Petrel (Oceanodromaleucorhda). -

Documentation of Recent Grand Banks Iceberg Grounding Events and a Comparison with Older Events of Known
Environmental Studies Research Funds 157 Documentation of Recent Iceberg Grounding Evens and a Comparison with Older Events of Know Age, Northern Grand Bank, Canada April 2006 Environmental Studies Research Funds Report ESRF # 157 June, 2005 DOCUMENTATION OF RECENT GRAND BANK ICEBERG GROUNDING EVENTS AND COMPARISON WITH OLDER EVENTS OF KNOWN AGE 1 1 2 3 Gary SonnichsenP ,P Thian HundertP ,P Robert Myers P ,P Patricia PocklingtonP P 1 P P Geological Survey of Canada (Atlantic), P.O. Box 1006, Dartmouth, Nova Scotia B2Y 4A2 [email protected] UTH 2 P P Rob Myers Consulting, 17 Harbour Drive, Dartmouth, Nova Scotia, B2Y 3N83 3 P P Arenicola Marine, Consultants in Marine Biology, 7 Bishop Ave., Wolfville N.S., B4P 2L3 The correct citation for this report is: Sonnichsen, G.V., Hundert, T., Pocklton, P. and Myers, R., 2005 Documentation of recent iceberg grounding events and a comparison with older events of known age, Northern Grand Bank, Canada, Environmental Studies Research Funds Report No.157, Calgary, 206 pp. The Environmental Studies Research Funds are financed from special levies on the oil and gas industry and administered by the National Energy Board for the Minister of Natural Resources Canada and the Minister of Indian Affairs and Northern Development. The Environmental Studies Research Funds and any person acting on their behalf assume no liability arising from the use of the information contained in this document. The opinions expressed are those of the authors and do not necessarily reflect those of the Environmental Studies Research Funds agencies. The use of trade names or identification of specific products does not constitute an endorsement or recommendation for use. -

International Black-Legged Kittiwake Conservation Strategy and Action Plan Acknowledgements Table of Contents
ARCTIC COUNCIL Circumpolar Seabird Expert Group July 2020 International Black-legged Kittiwake Conservation Strategy and Action Plan Acknowledgements Table of Contents Executive Summary ..............................................................................................................................................4 CAFF Designated Agencies: Chapter 1: Introduction .......................................................................................................................................5 • Norwegian Environment Agency, Trondheim, Norway Chapter 2: Ecology of the kittiwake ....................................................................................................................6 • Environment Canada, Ottawa, Canada Species information ...............................................................................................................................................................................................6 • Faroese Museum of Natural History, Tórshavn, Faroe Islands (Kingdom of Denmark) Habitat requirements ............................................................................................................................................................................................6 • Finnish Ministry of the Environment, Helsinki, Finland Life cycle and reproduction ................................................................................................................................................................................7 • Icelandic Institute of Natural -

Cursillo Parish Contacts
Anglican Diocese of Central Newfoundland Cursillo Parish Contacts Mailing Name Phone # Email Address Parish Address General Delivery Minnie Janes 536-3247 Badger’s Quay Badger’s Quay, NL A0G 1B0 POBox 942 545-2105 Edith Bagg [email protected] Bonavista Bonavista, NL A0C 1B0 470-0431 General Delivery Wilson & Stella Mills 656-4481 [email protected] Gander Bay Boyd’s Cove, NL A0G 1G0 POBox 59 Geraldine Purchase 672-3503 Buchans Buchans, NL A0H 1G0 POBox 45 June Holloway [email protected] Smith's Sound Port Blandford, NL A0C2G0 POBox 310 Rev. Terry Caines 891-1377 Burin Burin, NL A0G1E0 24 Park Avenue Elsie Sullivan 466-2002 [email protected] Clarenville Clarenville, NL A5A 1V8 POBox 111 Garry & Dallas Mitchell 884-5319 Twillingate Durrell, NL A0G1Y0 POBox 85 Gordon & Thelma Davidge 888-3336 [email protected] Belleoram English Harbour W, NL A0H 1M0 General Delivery Judy Mahoney Fogo Island Fogo, NL A0G 2B0 POBox 398 Jean Rose 832-2297 Fortune/Lamaline Fortune, NL A0E 1P0 POBox 391 Jean Eastman 674-5213 [email protected] Gambo Gambo, NL A0G 1T0 113 Ogilvie Street John & Beryl Barnes 256-8184 Gander Gander, NL A1V 2R2 POBox 24 Herbert & Beulah Ralph 533-2567 Glovertown Glovertown South, NL A0G 2M0 POBox 571 Winston & Shirley Walters 832-1930 [email protected] Grand Bank Grand Bank, NL A0E 1W0 20 Dunn Place Robert & Thelma Stockley 489-6945 [email protected] Grand Falls GrandFalls-Windsor, NL A2A2M3 8 Dorrity Place Ed & Glenda Warford 489-6747 [email protected] Windsor GrandFalls-Windsor, -

Feed Grain Transportation and Storage Assistance Regulations
CANADA CONSOLIDATION CODIFICATION Feed Grain Transportation and Règlement sur l’aide au Storage Assistance Regulations transport et à l’emmagasinage des céréales C.R.C., c. 1027 C.R.C., ch. 1027 Current to November 21, 2016 À jour au 21 novembre 2016 Published by the Minister of Justice at the following address: Publié par le ministre de la Justice à l’adresse suivante : http://laws-lois.justice.gc.ca http://lois-laws.justice.gc.ca OFFICIAL STATUS CARACTÈRE OFFICIEL OF CONSOLIDATIONS DES CODIFICATIONS Subsections 31(1) and (3) of the Legislation Revision and Les paragraphes 31(1) et (3) de la Loi sur la révision et la Consolidation Act, in force on June 1, 2009, provide as codification des textes législatifs, en vigueur le 1er juin follows: 2009, prévoient ce qui suit : Published consolidation is evidence Codifications comme élément de preuve 31 (1) Every copy of a consolidated statute or consolidated 31 (1) Tout exemplaire d'une loi codifiée ou d'un règlement regulation published by the Minister under this Act in either codifié, publié par le ministre en vertu de la présente loi sur print or electronic form is evidence of that statute or regula- support papier ou sur support électronique, fait foi de cette tion and of its contents and every copy purporting to be pub- loi ou de ce règlement et de son contenu. Tout exemplaire lished by the Minister is deemed to be so published, unless donné comme publié par le ministre est réputé avoir été ainsi the contrary is shown. publié, sauf preuve contraire. -
![2021 Special Collection Days[1] Copy](https://docslib.b-cdn.net/cover/3193/2021-special-collection-days-1-copy-903193.webp)
2021 Special Collection Days[1] Copy
2021 Special Collection Days Albert and Westmorland Counties from November 1st to April 30th on all roads opened by DOT and from May 1st to October 31st on all roads Communities Contractors Spring Summer Fall Hour* (unincorporated areas) Fero Waste & Albert Mines, Baltimore, Berryton, Caldedonia Recycling Inc. Mountain, Dawson Settlement, Edgetts Landing, May 12 August 4 October 6 Hillsborough West, Osborne Corner, Rosevale, 506-855-3376 Salem, Shenstone & Weldon Allison, Boundary Creek & Steeves Mountain June 2 August 18 October 20 Ammon, Irishtown, Lakeville (excluding Lakeside Estates & Weisner Rd), MacDougall Settlement, May 28 August 13 October 29 Notre-Dame (Westmorland County), Painsec, Saint- Philippe & Scotch Settlement Anagance (Westmorland County), Fawcett Hill, Glenvale (Westmorland County), Hillgrove, Intervale, June 4 August 20 October 22 Kinnear Settlement & Petitcodiac West Beaverbrook, Cape Enragé, Dennis Beach, Germantown, Harvey (Albert County), Hebron, May 10 August 2 October 4 Midway, New Horton, Waterside & West River Berry Mills, Lutes Mountain & Stilesville May 31 August 16 October 18 Calhoun, Greater Lakeburn, Meadowbrook, May 28 August 13 October 29 Scoudouc & Scoudouc Road 4:00 Canaan Station, Dundas (Westmorland County), AM Gallagher Ridge, Indian Mountain, McQuade & New June 1 August 17 October 19 Scotland Cape Station, Curryville, Hopewell Cape, Hopewell May 10 August 2 October 4 Hill, Lower Cape & Shepody (Albert County) Colpitts Settlement & Turtle Creek May 13 August 5 October 7 Dobson Corner, Harewood, -

Determining Fine-Scale Use and Movement Patterns of Diving Bird Species in Federal Waters of the Mid-Atlantic United States Using Satellite Telemetry
OCS Study BOEM 2017-069 Determining Fine-scale Use and Movement Patterns of Diving Bird Species in Federal Waters of the Mid-Atlantic United States Using Satellite Telemetry US Department of the Interior Bureau of Ocean Energy Management Office of Renewable Energy Programs OCS Study BOEM 2017-069 Determining Fine-scale Use and Movement Patterns of Diving Bird Species in Federal Waters of the Mid-Atlantic United States Using Satellite Telemetry Authors Caleb S. Spiegel, USFWS Division of Migratory Birds (Project Manager, Editor) Alicia M. Berlin, USGS Patuxent Wildlife Research Center Andrew T. Gilbert, Biodiversity Research Institute Carrie O. Gray, Biodiversity Research Institute William A. Montevecchi, Memorial University of Newfoundland Iain J. Stenhouse, Biodiversity Research Institute Scott L. Ford, Avian Specialty Veterinary Services Glenn H. Olsen, USGS Patuxent Wildlife Research Center Jonathan L. Fiely, USGS Patuxent Wildlife Research Center Lucas Savoy, Biodiversity Research Institute M. Wing Goodale, Biodiversity Research Institute Chantelle M. Burke, Memorial University of Newfoundland Prepared under BOEM Intra-agency Agreement #M12PG00005 by U.S. Department of Interior U.S. Fish and Wildlife Service Division of Migratory Birds 300 Westgate Center Dr. Hadley, MA 01035 Published by U.S. Department of the Interior Bureau of Ocean Energy Management Office of Renewable Energy Programs 2017-069 DISCLAIMER This study was funded by the US Department of the Interior, Bureau of Ocean Energy Management (BOEM), Environmental Studies Program, Washington, DC, through Intra-agency Agreement Number M12PG00005 with the US Department of Interior, US Fish and Wildlife Service, Division of Migratory Birds, Hadley, MA. This report has been technically reviewed by BOEM and it has been approved for publication.