Eukaryotic Cell Organelles
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

FIRST RECORD of Erythropsidinium Agile (GYMNODINIALES: WARNOWIACEAE) in the MEXICAN PACIFIC
CICIMAR Oceánides 25(2): 137-142 (2010) FIRST RECORD OF Erythropsidinium agile (GYMNODINIALES: WARNOWIACEAE) IN THE MEXICAN PACIFIC Primer registro de Erythropsidinium agile et Swezy, 1921, Proterythropsis Kofoid et Swezy, (Gymnodiniales: Warnowiaceae) en el 1921, Warnowia Lindemann, 1928, Greuetodinium Pacífico Mexicano Loeblich III, 1980, and Erythropsidinium P.C. Silva, 1960. Ten species of Erythropsidinium have been RESUMEN. Se registra por primera vez Erythropsi- described from warm and temperate seas. However, dinium agile, un dinoflagelado de la Familia Warno- a taxonomical study based on the changes in struc- wiaceae para el Pacífico Mexicano, dentro de Bahía ture, position, and coloration of the ocelloid in the de La Paz (Golfo de California). Se observaron 26 course of the cell division or individual development ejemplares de E. agile, principalmente en muestras revealed that some species had different morpho- de fitoplancton de red para el periodo de estudio (Ju- types (Elbrächter, 1979). At present the valid species nio, 2006 a Junio, 2010). En muestras de botella se currently considered to belong to this genus are: estimaron densidades entre 80 y 1000 cél. L–1. Los ejemplares de E. agile mostraron gran variación en E. agile (Hertwig, 1884) P.C. Silva, 1960, E. cochlea forma, tamaño y coloración; se presentaron princi- (Schütt, 1895) P.C. Silva, 1960, E. extrudens (Ko- palmente en el período invierno-primavera, cuando foid et Swezy, 1921) P.C. Silva, 1960, and E. minus la columna del agua está homogénea, a temperatu- (Kofoid et Swezy, 1921) P.C. Silva, 1960. For the ras entre 19 y 22 °C y rica en nutrientes. -

Download the Abstract Book
1 Exploring the male-induced female reproduction of Schistosoma mansoni in a novel medium Jipeng Wang1, Rui Chen1, James Collins1 1) UT Southwestern Medical Center. Schistosomiasis is a neglected tropical disease caused by schistosome parasites that infect over 200 million people. The prodigious egg output of these parasites is the sole driver of pathology due to infection. Female schistosomes rely on continuous pairing with male worms to fuel the maturation of their reproductive organs, yet our understanding of their sexual reproduction is limited because egg production is not sustained for more than a few days in vitro. Here, we explore the process of male-stimulated female maturation in our newly developed ABC169 medium and demonstrate that physical contact with a male worm, and not insemination, is sufficient to induce female development and the production of viable parthenogenetic haploid embryos. By performing an RNAi screen for genes whose expression was enriched in the female reproductive organs, we identify a single nuclear hormone receptor that is required for differentiation and maturation of germ line stem cells in female gonad. Furthermore, we screen genes in non-reproductive tissues that maybe involved in mediating cell signaling during the male-female interplay and identify a transcription factor gli1 whose knockdown prevents male worms from inducing the female sexual maturation while having no effect on male:female pairing. Using RNA-seq, we characterize the gene expression changes of male worms after gli1 knockdown as well as the female transcriptomic changes after pairing with gli1-knockdown males. We are currently exploring the downstream genes of this transcription factor that may mediate the male stimulus associated with pairing. -

Autophagy in Trypanosomatids
Cells 2012, 1, 346-371; doi:10.3390/cells1030346 OPEN ACCESS cells ISSN 2073-4409 www.mdpi.com/journal/cells Review Autophagy in Trypanosomatids Ana Brennand 1,†, Eva Rico 2,†,‡ and Paul A. M. Michels 1,* 1 Research Unit for Tropical Diseases, de Duve Institute, Université catholique de Louvain, Avenue Hippocrate 74, postal box B1.74.01, B-1200 Brussels, Belgium; E-Mail: [email protected] 2 Department of Biochemistry and Molecular Biology, University Campus, University of Alcalá, Alcalá de Henares, Madrid, 28871, Spain; E-Mail: [email protected] † These authors contributed equally to this work. ‡ Present Address: Centre for Immunity, Infection and Evolution, Institute of Immunology and Infection Research, School of Biological Sciences, King’s Buildings, University of Edinburgh, West Mains Road, Edinburgh EH9 3JT, UK. * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel.: +32-2-7647473; Fax: +32-2-7626853. Received: 28 June 2012; in revised form: 14 July 2012 / Accepted: 16 July 2012 / Published: 27 July 2012 Abstract: Autophagy is a ubiquitous eukaryotic process that also occurs in trypanosomatid parasites, protist organisms belonging to the supergroup Excavata, distinct from the supergroup Opistokontha that includes mammals and fungi. Half of the known yeast and mammalian AuTophaGy (ATG) proteins were detected in trypanosomatids, although with low sequence conservation. Trypanosomatids such as Trypanosoma brucei, Trypanosoma cruzi and Leishmania spp. are responsible for serious tropical diseases in humans. The parasites are transmitted by insects and, consequently, have a complicated life cycle during which they undergo dramatic morphological and metabolic transformations to adapt to the different environments. -
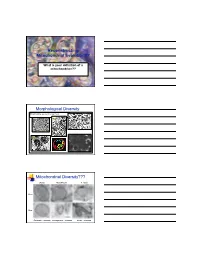
Reconstructing Mitochondrial Evolution?? Morphological Diversity Mitochondrial Diversity???
Reconstructing Mitochondrial Evolution?? What is your definition of a mitochondrion?? Morphological Diversity Mitochondria as we all know them: Suprarenal gland Liver cell Plasma cell Adrenal cortex Mitochondrial Diversity??? Chicken Neocallimastix T. foetus 100 nm 50 nm Entamoeba - mitosome microsporidian - mitosome Giardia - mitosome Reconstructing Evolution Mitochondrial evolution well established endosymbiotic theory α-proteobacterium - Rickettsia prowazekii Hydrogenosomal evolution No DNA NOW 2 examples Nyctotherus and Blastocystis (MLO) Several proteins similar to mitochondria Mitosome evolution No DNA Few proteins identified similar to mitochondria Origins via Endosymbiosis Current dogma - mitochondria and related Aerobic α-proteobacterium prokaryote gave rise to present day mitochondria. organelles arose just once in evolution Are hydrogenosomes and mitosomes of anaerobic protists derived from the same proto-mitochondrion? Evidence for: accumulating evidence for several proteins that are currently found in mitochondria - Proteins of Fe-S cluster formation. Scenario A Common ancestor facultative Scenario B organism? degenerate mitochondrion invoke lateral gene transfer from anaerobic prokaryotes Hypotheses for Mito Acquisition Mitochondrion-related Organelles 2 way to interpret this summary - can you think of both? Groups with mitochondrial homologues Hjort et al Phil. Trans. R. Soc. B 2010 365, 713-727 Origin of Hydrogenosomes (Mitosomes?) 1. a)Conversion of Mitochondria HYD b)Common ancestor with mitochondria ? HYD 2. Independent Origin α-proteobacterium ? - anaerobic bacterium HYD Organelles - origins and biogenesis Approaches: (1) Conduct phylogenetic analyses of similar proteins Hsp70 Fd Hsp60 Isc subunits (2) Examine protein targeting to the organelle matrix protein targeting membrane protein targeting (3) Characterize membrane/translocation components These components could have evolved as the endosymbiont was converted to organelle. Reveals evolutionary history. -

The Cytological Events and Molecular Control of Life Cycle Development of Trypanosoma Brucei in the Mammalian Bloodstream
pathogens Review The Cytological Events and Molecular Control of Life Cycle Development of Trypanosoma brucei in the Mammalian Bloodstream Eleanor Silvester †, Kirsty R. McWilliam † and Keith R. Matthews * Institute for Immunology and Infection Research, Centre for Immunity, Infection and Evolution, School of Biological Sciences, King’s Buildings, University of Edinburgh, Charlotte Auerbach Road, Edinburgh EH9 3FL, UK; [email protected] (E.S.); [email protected] (K.R.McW.) * Correspondence: [email protected]; Tel.: +44-131-651-3639 † These authors contributed equally to this work. Received: 23 May 2017; Accepted: 22 June 2017; Published: 28 June 2017 Abstract: African trypanosomes cause devastating disease in sub-Saharan Africa in humans and livestock. The parasite lives extracellularly within the bloodstream of mammalian hosts and is transmitted by blood-feeding tsetse flies. In the blood, trypanosomes exhibit two developmental forms: the slender form and the stumpy form. The slender form proliferates in the bloodstream, establishes the parasite numbers and avoids host immunity through antigenic variation. The stumpy form, in contrast, is non-proliferative and is adapted for transmission. Here, we overview the features of slender and stumpy form parasites in terms of their cytological and molecular characteristics and discuss how these contribute to their distinct biological functions. Thereafter, we describe the technical developments that have enabled recent discoveries that uncover how the slender to stumpy transition is enacted in molecular terms. Finally, we highlight new understanding of how control of the balance between slender and stumpy form parasites interfaces with other components of the infection dynamic of trypanosomes in their mammalian hosts. -

Kmcb-Abstract-Book-2019-Final
VIII April 27 – May 1, 2019 2 Eight Kinetoplastid Molecular Cell Biology Meeting April 27 – May 1, 2019 Hosted by the Marine Biological Laboratory Woods Hole, Massachusetts, USA The meeting was founded by George A.M. Cross in 2005 Organizers George A.M. Cross 2005-2011 Christian Tschudi 2013-2019 3 KMCBM 2019 Acknowledgements The organizer wishes to thank: The Program Committee Barbara Burleigh (Harvard T. H. Chan School of Public Health, Boston, USA) James D. Bangs (University at Buffalo, Buffalo, USA) Stephen M. Beverley (Washington University School of Medicine, St. Louis, USA) Keith R. Matthews (The University of Edinburgh, Edinburgh, Scotland, UK) The Staff at MBL: Paul Anderson and the MBL Housing and Conference Staff for registration and housing; All the IT AV Support staff and the staff in Sodexo Food Service at the MBL. Cover Design: Markus Engstler 4 KMCBM 2019 Program Saturday, April 27 02:00 – 05:00 Arrival, Registration and Poster Session A setup 04:00 – 06:30 Greeting and Dinner 07:00 – 09:00 Session I: VSG (chair: Mark Carrington) 09:00 – 11:00 Mixer Sunday, April 28 07:00 – 08:30 Breakfast 08:45 – 11:45 Session II: Biochemistry/Metabolism (chair: Ken Stuart) 12:00 – 01:30 Lunch 02:00 – 04:30 Session III: Cell Biology (chair: Kimberly Paul) 06:00 – 07:00 Dinner 07:00 – 09:00 POSTER PRESENTATIONS: Session A 09:00 – 11:00 Mixer & Poster A/B Changeover Monday, April 29 07:00 – 08:30 Breakfast 08:45 – 11:45 Session IV: Pathogenesis I (chair: Luisa Figueiredo) 12:00 – 01:30 Lunch 01:30 – 06:00 Free Time 06:00 – 07:00 Dinner 07:00 -

Characterising Planktonic Dinoflagellate Diversity in Singapore Using DNA Metabarcoding
Metabarcoding and Metagenomics 2: 1–14 DOI 10.3897/mbmg.2.25136 Research Article Characterising planktonic dinoflagellate diversity in Singapore using DNA metabarcoding Yue Sze1, Lilibeth N. Miranda2, Tsai Min Sin2,†, Danwei Huang1,2 1 Department of Biological Sciences, National University of Singapore, Singapore 117558, Singapore. 2 Tropical Marine Science Institute, National University of Singapore, Singapore 119227, Singapore. † Deceased. Corresponding author: Danwei Huang ([email protected]) Academic editor: Thorsten Stoeck | Received 19 March 2018 | Accepted 24 April 2018 | Published 17 May 2018 Abstract Dinoflagellates are traditionally identified morphologically using microscopy, which is a time-consuming and labour-intensive process. Hence, we explored DNA metabarcoding using high-throughput sequencing as a more efficient way to study planktonic dinoflagellate diversity in Singapore’s waters. From 29 minimally pre-sorted water samples collected at four locations in western Singapore, DNA was extracted, amplified and sequenced for a 313-bp fragment of the V4–V5 region in the 18S ribosomal RNA gene. Two sequencing runs generated 2,847,170 assembled paired-end reads, corresponding to 573,176 unique sequences. Sequenc- es were clustered at 97% similarity and analysed with stringent thresholds (≥150 bp, ≥20 reads, ≥95% match to dinoflagellates), recovering 28 dinoflagellate taxa. Dinoflagellate diversity captured includes parasitic and symbiotic groups which are difficult to identify morphologically. Richness is similar between the inner and outer West Johor Strait, but variations in community structure are apparent, likely driven by environmental differences. None of the taxa detected in a recent phytoplankton bloom along the West Johor Strait have been recovered in our samples, suggesting that background communities are distinct from bloom communities. -

The Revised Classification of Eukaryotes
See discussions, stats, and author profiles for this publication at: https://www.researchgate.net/publication/231610049 The Revised Classification of Eukaryotes Article in Journal of Eukaryotic Microbiology · September 2012 DOI: 10.1111/j.1550-7408.2012.00644.x · Source: PubMed CITATIONS READS 961 2,825 25 authors, including: Sina M Adl Alastair Simpson University of Saskatchewan Dalhousie University 118 PUBLICATIONS 8,522 CITATIONS 264 PUBLICATIONS 10,739 CITATIONS SEE PROFILE SEE PROFILE Christopher E Lane David Bass University of Rhode Island Natural History Museum, London 82 PUBLICATIONS 6,233 CITATIONS 464 PUBLICATIONS 7,765 CITATIONS SEE PROFILE SEE PROFILE Some of the authors of this publication are also working on these related projects: Biodiversity and ecology of soil taste amoeba View project Predator control of diversity View project All content following this page was uploaded by Smirnov Alexey on 25 October 2017. The user has requested enhancement of the downloaded file. The Journal of Published by the International Society of Eukaryotic Microbiology Protistologists J. Eukaryot. Microbiol., 59(5), 2012 pp. 429–493 © 2012 The Author(s) Journal of Eukaryotic Microbiology © 2012 International Society of Protistologists DOI: 10.1111/j.1550-7408.2012.00644.x The Revised Classification of Eukaryotes SINA M. ADL,a,b ALASTAIR G. B. SIMPSON,b CHRISTOPHER E. LANE,c JULIUS LUKESˇ,d DAVID BASS,e SAMUEL S. BOWSER,f MATTHEW W. BROWN,g FABIEN BURKI,h MICAH DUNTHORN,i VLADIMIR HAMPL,j AARON HEISS,b MONA HOPPENRATH,k ENRIQUE LARA,l LINE LE GALL,m DENIS H. LYNN,n,1 HILARY MCMANUS,o EDWARD A. D. -

Septal Pore Caps in Basidiomycetes Composition and Ultrastructure
Septal Pore Caps in Basidiomycetes Composition and Ultrastructure Septal Pore Caps in Basidiomycetes Composition and Ultrastructure Septumporie-kappen in Basidiomyceten Samenstelling en Ultrastructuur (met een samenvatting in het Nederlands) Proefschrift ter verkrijging van de graad van doctor aan de Universiteit Utrecht op gezag van de rector magnificus, prof.dr. J.C. Stoof, ingevolge het besluit van het college voor promoties in het openbaar te verdedigen op maandag 17 december 2007 des middags te 16.15 uur door Kenneth Gregory Anthony van Driel geboren op 31 oktober 1975 te Terneuzen Promotoren: Prof. dr. A.J. Verkleij Prof. dr. H.A.B. Wösten Co-promotoren: Dr. T. Boekhout Dr. W.H. Müller voor mijn ouders Cover design by Danny Nooren. Scanning electron micrographs of septal pore caps of Rhizoctonia solani made by Wally Müller. Printed at Ponsen & Looijen b.v., Wageningen, The Netherlands. ISBN 978-90-6464-191-6 CONTENTS Chapter 1 General Introduction 9 Chapter 2 Septal Pore Complex Morphology in the Agaricomycotina 27 (Basidiomycota) with Emphasis on the Cantharellales and Hymenochaetales Chapter 3 Laser Microdissection of Fungal Septa as Visualized by 63 Scanning Electron Microscopy Chapter 4 Enrichment of Perforate Septal Pore Caps from the 79 Basidiomycetous Fungus Rhizoctonia solani by Combined Use of French Press, Isopycnic Centrifugation, and Triton X-100 Chapter 5 SPC18, a Novel Septal Pore Cap Protein of Rhizoctonia 95 solani Residing in Septal Pore Caps and Pore-plugs Chapter 6 Summary and General Discussion 113 Samenvatting 123 Nawoord 129 List of Publications 131 Curriculum vitae 133 Chapter 1 General Introduction Kenneth G.A. van Driel*, Arend F. -

Mitosomes in Entamoeba Histolytica Contain a Sulfate Activation Pathway
Mitosomes in Entamoeba histolytica contain a sulfate activation pathway Fumika Mi-ichi, Mohammad Abu Yousuf, Kumiko Nakada-Tsukui, and Tomoyoshi Nozaki1 Department of Parasitology, National Institute of Infectious Diseases, Tokyo 162-8640, Japan Edited by Andrew Roger, Dalhousie University, Halifax, NS, Canada, and accepted by the Editorial Board October 19, 2009 (received for review June 25, 2009) Hydrogenosomes and mitosomes are mitochondrion-related or- and M. balamuthi lack the ISC system, and instead possess the ganelles in anaerobic/microaerophilic eukaryotes with highly re- nitrogen fixation (NIF) system, which is most likely derived from an duced and divergent functions. The full diversity of their content ancestral nitrogen fixing -proteobacterium by lateral gene transfer and function, however, has not been fully determined. To under- (22, 24). Only 5 proteins have been demonstrated in E. histolytica stand the central role of mitosomes in Entamoeba histolytica,a mitosomes: Cpn60 (8–10, 12), Cpn10 (13), mitochondrial Hsp70 parasitic protozoon that causes amoebic dysentery and liver ab- (11, 15), pyridine nucleotide transhydrogenase (PNT) (2, 8), and scesses, we examined the proteomic profile of purified mitosomes. mitochondria carrier family (MCF, ADP/ATP transporter) (14), Using 2 discontinuous Percoll gradient centrifugation and MS and the central role of mitosomes in E. histolytica remains unknown. analysis, we identified 95 putative mitosomal proteins. Immuno- Analysis of the genome of E. histolytica has not revealed any fluorescence assay showed that 3 proteins involved in sulfate additional information regarding the function of mitosomes and activation, ATP sulfurylase, APS kinase, and inorganic pyrophos- thus, a proteomic analysis of mitosomes seems to be the best phatase, as well as sodium/sulfate symporter, involved in sulfate approach to understand its structure and function (1, 2). -

A Novel ADP/ATP Transporter in the Mitosome of the Microaerophilic Human Parasite Entamoeba Histolytica
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Elsevier - Publisher Connector Current Biology, Vol. 15, 737–742, April 26, 2005, ©2005 Elsevier Ltd All rights reserved. DOI 10.1016/j.cub.2005.02.068 A Novel ADP/ATP Transporter in the Mitosome of the Microaerophilic Human Parasite Entamoeba histolytica Ka Wai Chan,1,5 Dirk-Jan Slotboom,1,5 Sian Cox,3 ses confirm that the Entamoeba ADP/ATP carrier is T. Martin Embley,2,* Olivier Fabre,1 distinct from archetypal mitochondrial ADP/ATP carri- Mark van der Giezen,3,6 Marilyn Harding,1 ers, an observation that is supported by its different David S. Horner,4 Edmund R.S. Kunji,1,* substrate and inhibitor specificity. Because many Gloria León-Avila,3,7 and Jorge Tovar3 functions of yeast and human mitochondria rely on 1Dunn Human Nutrition Unit solutes transported by specialized members of this Medical Research Council family, the Entamoeba mitosome must contain only Hills Road a small subset of these processes requiring adenine Cambridge CB2 2XY nucleotide exchange. United Kingdom 2 School of Biology The Devonshire Building Results and Discussion University of Newcastle upon Tyne Newcastle NE1 7RU Identification of an MCF Homolog United Kingdom on the Entamoeba histolytica Genome 3 School of Biological Sciences We identified a homolog of the mitochondrial carrier Royal Holloway family (MCF) on the Entamoeba histolytica genome (TIGR) University of London with BLAST searches. The translated protein contains a Egham, Surrey TW20 0EX tripartite structure [8], signature motifs (IPR001993), and United Kingdom other features typical of MCF members (Figure 1A). -

Effect of Hydroxyurea on the Intracellular Multiplication of Toxoplasma Gondii, Leishmania Amazonensis and Trypanosoma Cruzi
Brazilian Journal of Medical and Biological Research (2003) 36: 65-69 Hydroxyurea inhibits intracellular parasites 65 ISSN 0100-879X Short Communication Effect of hydroxyurea on the intracellular multiplication of Toxoplasma gondii, Leishmania amazonensis and Trypanosoma cruzi E.J.T. Melo and Laboratório de Biologia Celular, Centro de Biociências e Biotecnologia, H.J. Beiral Universidade Estadual do Norte Fluminense, Campos dos Goytacazes, RJ, Brasil Abstract Correspondence Toxoplasma gondii, Leishmania amazonensis and Trypanosoma Key words E.J.T. Melo cruzi are obligate intracellular parasites that multiply until lysis of host · Toxoplasma gondii Laboratório de Biologia Celular cells. The present study was undertaken to evaluate the effect of · Trypanosoma cruzi CBB, Universidade Estadual do hydroxyurea (an inhibitor of cell division at the G1/S phase) on the · Leishmania amazonensis Norte Fluminense multiplication of L. amazonensis, T. gondii, and T. cruzi in infected · Hydroxyurea Av. Alberto Lamego, 2000 host cells. Infected cells were treated with hydroxyurea (4 mM) for 48 28013-600 Campos dos Goytacazes, RJ h. Hydroxyurea arrested intracellular multiplication of all infective Brasil Fax: +55-022-2726-1514 forms of the parasites tested. In treated cultures, the percent of infected E-mail: [email protected] host cells decreased (50-97%) and most intracellular parasites were eliminated. Ultrastructural observations showed no morphologic change in host cells while intracellular parasites presented drastic morpho- logic alterations or disruption. The results strongly suggest that hy- Received May 29, 2001 droxyurea was able to interfere with the multiplication of intracellular Accepted October 14, 2002 parasites, leading to an irreversible morphological effect on L. amazo- nensis, T. gondii, and T.