Precision of Helicopter-Based Total-Area Counts of Large Ungulates in Bushveld
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
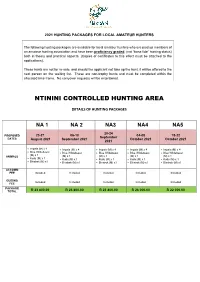
Ntinini Controlled Hunting Area
2021 HUNTING PACKAGES FOR LOCAL AMATEUR HUNTERS The following hunting packages are available for local amateur hunters who are paid up members of an amateur hunting association and have been proficiency graded, (not “bona fide” hunting status) both in theory and practical aspects. (Copies of certificates to this effect must be attached to the applications). These hunts are not for re-sale, and should the applicant not take up the hunt, it will be offered to the next person on the waiting list. These are non-trophy hunts and must be completed within the allocated time frame. No carryover requests will be entertained. NTININI CONTROLLED HUNTING AREA DETAILS OF HUNTING PACKAGES NA 1 NA 2 NA3 NA4 NA5 20-24 23-27 06-10 04-08 18-22 PROPOSED September DATES August 2021 September 2021 October 2021 October 2021 2021 • Impala (M) x 4 • Impala (M) x 4 • Impala (M) x 4 • Impala (M) x 4 • Impala (M) x 4 • Blue Wildebeest • Blue Wildebeest • Blue Wildebeest • Blue Wildebeest • Blue Wildebeest (M) x 1 ANIMALS (M) x 1 (M) x 1 (M) x 1 (M) x 1 • Kudu (M) x 1 • Kudu (M) x 1 • Kudu (M) x 1 • Kudu (M) x 1 • Kudu (M) x 1 • Blesbok (M) x1 • Blesbok (M) x1 • Blesbok (M) x1 • Blesbok (M) x1 • Blesbok (M) x1 ACCOMM FEE Included Included Included Included Included GUIDING Included Included Included Included Included FEE PACKAGE TOTAL R 23 800.00 R 23 800.00 R 23 800.00 R 26 000.00 R 22 000.00 ITHALA CONTROLLED HUNTING AREA DETAILS OF HUNTING PACKAGES IGRA 1 IGRA 2 IGRA 3 IGRA 4 PROPOSED 11-16 02-07 16-21 06-11 DATES June 2021 July 2021 July 2021 August 2021 • Impala -

Pending World Record Waterbuck Wins Top Honor SC Life Member Susan Stout Has in THIS ISSUE Dbeen Awarded the President’S Cup Letter from the President
DSC NEWSLETTER VOLUME 32,Camp ISSUE 5 TalkJUNE 2019 Pending World Record Waterbuck Wins Top Honor SC Life Member Susan Stout has IN THIS ISSUE Dbeen awarded the President’s Cup Letter from the President .....................1 for her pending world record East African DSC Foundation .....................................2 Defassa Waterbuck. Awards Night Results ...........................4 DSC’s April Monthly Meeting brings Industry News ........................................8 members together to celebrate the annual Chapter News .........................................9 Trophy and Photo Award presentation. Capstick Award ....................................10 This year, there were over 150 entries for Dove Hunt ..............................................12 the Trophy Awards, spanning 22 countries Obituary ..................................................14 and almost 100 different species. Membership Drive ...............................14 As photos of all the entries played Kid Fish ....................................................16 during cocktail hour, the room was Wine Pairing Dinner ............................16 abuzz with stories of all the incredible Traveler’s Advisory ..............................17 adventures experienced – ibex in Spain, Hotel Block for Heritage ....................19 scenic helicopter rides over the Northwest Big Bore Shoot .....................................20 Territories, puku in Zambia. CIC International Conference ..........22 In determining the winners, the judges DSC Publications Update -

Influence of Common Eland (Taurotragus Oryx) Meat Composition on Its Further Technological Processing
CZECH UNIVERSITY OF LIFE SCIENCES PRAGUE Faculty of Tropical AgriSciences Department of Animal Science and Food Processing Influence of Common Eland (Taurotragus oryx) Meat Composition on its further Technological Processing DISSERTATION THESIS Prague 2018 Author: Supervisor: Ing. et Ing. Petr Kolbábek prof. MVDr. Daniela Lukešová, CSc. Co-supervisors: Ing. Radim Kotrba, Ph.D. Ing. Ludmila Prokůpková, Ph.D. Declaration I hereby declare that I have done this thesis entitled “Influence of Common Eland (Taurotragus oryx) Meat Composition on its further Technological Processing” independently, all texts in this thesis are original, and all the sources have been quoted and acknowledged by means of complete references and according to Citation rules of the FTA. In Prague 5th October 2018 ………..………………… Acknowledgements I would like to express my deep gratitude to prof. MVDr. Daniela Lukešová CSc., Ing. Radim Kotrba, Ph.D. and Ing. Ludmila Prokůpková, Ph.D., and doc. Ing. Lenka Kouřimská, Ph.D., my research supervisors, for their patient guidance, enthusiastic encouragement and useful critiques of this research work. I am very gratefull to Ing. Petra Maxová and Ing. Eva Kůtová for their valuable help during the research. I am also gratefull to Mr. Petr Beluš, who works as a keeper of elands in Lány, Mrs. Blanka Dvořáková, technician in the laboratory of meat science. My deep acknowledgement belongs to Ing. Radek Stibor and Mr. Josef Hora, skilled butchers from the slaughterhouse in Prague – Uhříněves and to JUDr. Pavel Jirkovský, expert marksman, who shot the animals. I am very gratefull to the experts from the Natura Food Additives, joint-stock company and from the Alimpex-maso, Inc. -

South Africa Log Illustrated
South Africa 11 November - 1 December, 2016 (CIA Map) South Africa - a land I knew only from a map. Africa - a continent I’d never visited. This trip is one of a life time. And so it begins. Departing 11-12 November, Friday-Saturday Typical. Leaving NY turned out to be an ordeal. The plane change at JFK allowed only an hour between Jet Blue landing and South African airways boarding, which included changing terminals in a strange airport, and going through security again - hardly enough time at an empty airport, much less at one of the busiest in the world. Spending the night in Boston and getting an earlier Jet Blue flight was in order. I went to Logan the night before to change the flight, which is where the ordeal begins - waiting at Jet Blue full service desk, then more waiting while the agent called higher ups to see about changing, then good news that the flight could be changed, then bad news - because it was a South African Airways ticket, Jet Blue couldn’t do it, !1 then more waiting to check again, then good news that it was all set, then more waiting for the ticket to be into the system to print the boarding pass, then more waiting when it didn’t happen, then more waiting to recall and check, then more waiting to determine it couldn’t be done (can only change tickets on the day of the flight, not within 24 hours!), then bad news because my original ticket had been cancelled, then more waiting to call South African airways to reissue the boarding pass, then more waiting to refund the $50 for a ticket change. -

A National Park in Northern Rhodesia 15
A National Park in Northern Rhodesia 15 A NATIONAL PARK IN NORTHERN RHODESIA By a Proclamation dated 20th April, 1950, the Governor of Northern Rhodesia has established a National Park, to be known as the Kafue National Park. This new park covers some 8,650 square miles roughly in the central Kafue basin, between latitudes 14° and 16° 40' S. It contains a wide range of country from the comparatively drv sandy lands of the south to the big rivers, swamps, and heavy timber of the northern section. The magnificent Kafue River dominates the whole central portion, adding scenic beauty to the attraction of wild life. The park contains representatives of most species of the fauna of Northern Rhodesia. Primates are represented by the Rhodesian Baboon, the Vervet Monkey, and the Greater and Lesser Night-Apes. There are Elephant and Black Rhinoceros, Buffalo, Lion, Leopard, Cheetah, and numbers of the smaller carnivora. Antelope include Eland, Sable, Roan, Liehtenstein's Hartebeest, Blue Wildebeest, Kudu, Defassa Waterbuck, Bushbuck, Reedbuck, Puku, Impala, Oribi, Common, Blue, and Yellow-backed Duikers, Klipspringer, and Sharpe's Stein- buck. There are Red Lechwe and Sitatunga in the Busango Swamp in the north, Hippopotamus in numbers in the Kafue and its larger tributaries. Zebra are common, Warthog and Bushpig everywhere. Birds are abundant. The park is uninhabited apart from certain small settle- ments, on a limited section of the Kafue River, belonging to the indigenous Africans under their tribal chief Kayingu. These people must be accorded their traditional local hunting rights, but such will affect only a small fraction of the whole vast wild area. -

Parasites of South African Wildlife. XIX. the Prevalence of Helminths in Some Common Antelopes, Warthogs and a Bushpig in the Limpopo Province, South Africa
Page 1 of 11 Original Research Parasites of South African wildlife. XIX. The prevalence of helminths in some common antelopes, warthogs and a bushpig in the Limpopo province, South Africa Authors: Little work has been conducted on the helminth parasites of artiodactylids in the northern 1 Ilana C. van Wyk and western parts of the Limpopo province, which is considerably drier than the rest of the Joop Boomker1 province. The aim of this study was to determine the kinds and numbers of helminth that Affiliations: occur in different wildlife hosts in the area as well as whether any zoonotic helminths were 1Department of Veterinary present. Ten impalas (Aepyceros melampus), eight kudus (Tragelaphus strepsiceros), four blue Tropical Diseases, University wildebeest (Connochaetes taurinus), two black wildebeest (Connochaetes gnou), three gemsbok of Pretoria, South Africa (Oryx gazella), one nyala (Tragelaphus angasii), one bushbuck (Tragelaphus scriptus), one Correspondence to: waterbuck (Kobus ellipsiprymnus), six warthogs (Phacochoerus aethiopicus) and a single bushpig Ilana van Wyk (Potamochoerus porcus) were sampled from various localities in the semi-arid northern and western areas of the Limpopo province. Email: [email protected] New host–parasite associations included Trichostrongylus deflexus from blue wildebeest, Postal address: Agriostomum gorgonis from black wildebeest, Stilesia globipunctata from the waterbuck and Private bag X04, Fasciola hepatica in a kudu. The mean helminth burden, including extra-gastrointestinal Onderstepoort 0110, South Africa helminths, was 592 in impalas, 407 in kudus and blue wildebeest, 588 in black wildebeest, 184 in gemsbok, and 2150 in the waterbuck. Excluding Probstmayria vivipara, the mean helminth Dates: burden in warthogs was 2228 and the total nematode burden in the bushpig was 80. -

South Africa Travel Guide 2017
South Africa Travel Guide 2017 1 From the Editor... After a few failed attempts at collecting travel information about South Africa, I decided it would be a great idea to publish my own South Africa Travel Guide. It has taken me about 3 years to assemble this valuable publication (in between extra hours in the CLO Office and publishing JJ’s and Classifieds, and more Classi- fieds, and more JJ’s). Realistically, I thought I would lose my mind if I heard, “I will send over travel brochures ‘just now’” one more time... It has been a lot of work, but being in the CLO Office is the reason that I started this venture in the first place. My favorite part of working in the CLO Office is helping people who are searching for travel information. There is no greater reward as the Editor of the Jacaranda Journal, than to hear that one of my readers has booked a vacation or some sort of adventure because of a travel story or advice from our office. Travelling means taking a break from everyday routines and just enjoying life. I personally believe that there is so much benefit to travel, which is why I am hoping this Guide entices you to travel more. Travel gives us better perspective, it makes us more adaptable and adventurous, and it just makes people happy. We are in a unique position, living life in the Foreign Service, and one of the greatest benefits is seeing the world. We get the opportunity to see places we would never have dreamed of and even better, we get to share them sometimes with friends and family. -

SOUTH AFRICA: FAIREST CAPE to KRUGER 7Th – 30Th January 2016
Tropical Birding Trip Report South Africa Set Departure: January 2016 A Tropical Birding SET DEPARTURE tour SOUTH AFRICA: FAIREST CAPE TO KRUGER th th 7 – 30 January 2016 Karoo Korhaan is a raucous endemic we encounter frequently in the Karoo and Agulhas parts of this tour Tour Leader: Lisle Gwynn All photos in this report were taken by Lisle Gwynn. Species pictured are highlighted RED. 1 www.tropicalbirding.com +1-409-515-0514 [email protected] Page Tropical Birding Trip Report South Africa Set Departure: January 2016 INTRODUCTION South Africa is a land of many contrasts and incredible natural wealth, and the set departure tour is designed to take in as much of this as possible. From the fresh breezes and roaring ocean of the Cape to the dry savannah of the Kruger National Park, and down to the lush green montane forests of KwaZulu-Natal, we bird our way through many different habitat types and go in search of the myriad birds that call this place home, be it for the whole year or just part of it. The tour followed our tried and tested route through the roaring but san-washed Cape and along the south coast into the Garden Route. From there we moved inland to the unique dry landscapes of the Karoo and Tankwa Karoo before flying up-country to Johannesburg and exploring the mid-altitude grasslands of Wakkerstroom. After a few days in the world famous Kruger National Park we then backtracked to Jo’burg before winding our way through Zululand, visiting Mkhuze Game Reserve, the St. -

Engl South Africa Limpopo, Game Reserves & Brandberg 2017
South Africa 2017 - Limpopo , Game Reserve s & Brandberg - Office Germany: Office Austria: Ziegelstadel 1 · D-88316 Isny Europastrasse 1/1 · A-7540 Güssing Phone.: +49 (0) 75 62 / 914 54 - 14 Phone: +43 (0) 33 22 / 42 963 - 0 www.blaser-safaris.com Fax.: +43 (0) 33 22 / 42 963 - 59 [email protected] Hunt in South In South Africa, safaris can be conducted throughout the year; however the best time Africa : for a safari is between April & October. Some 40 different species of game can be hunted across the country, but each safari is individually planned to give you maximum enjoyment & satisfaction. The safari is suitable for the whole family, for non-hunters; there are a variety of inter- esting, photographic tours. All wildlife species listed in the price list are hunted on farmland / hunting areas around the lodge. LIMPOPO Limpopo The area of our partner Wayne Wagner Safaris is located in Hoedspruit Limpopo Pro v- ince and is situated between Phalaborwa, Gravelotte and Mica on the Olifants River. Accommodation To be sure that the safari meets with your specific requirements a questionnaire will be for Limpopo & completed before your arrival with your personal details, species to be hunted, food Games Reserves preferences and accommodation requirements. - 2 - Prices 201 7 5 DAY SOUTH AFRICAN PLAINS GAME CULLING PACKAGE The following package is a cull hunt for certain plains game species. This package includes 5 full days of hunting; you must plan on arriving one day prior to commencement of the safari. The hunt will take place in the bushveld region of the Limpopo Province near the town of Hoedspruit. -

The Effect of Species Associations on the Diversity and Coexistence of African Ungulates
The effect of species associations on the diversity and coexistence of African ungulates. By Nancy Barker For Professor Kolasa BIO306H1 – Tropical Ecology University of Toronto Wednesday, August 24th, 2005 Abstract: The effects of species associations on species diversity and coexistence were investigated in East Africa. The frequency and group sizes of African ungulates were observed and analyzed to determine for differences in species associations based on their density and distribution, as well as their associations with other species. Associations between species were determined to be nonrandom and seen to affect the demographics of associating herds. Such associations mirrored in other studies were shown to be the result of interspecific competition, habitat preferences and predation pressure which increases the potential for coexistence between species. This suggests a potentially important role in the regulation of species diversity by ecological dynamics in species rich communities. In the face of today’s biodiversity crisis, such understanding of species associations and how they are regulated may have huge implications for conservation. Introduction: known with the famous Darwin’s finches of the Galapagos Islands. However, there are many other Sympatric coexistence of organisms within a guilds with what seems to be extensive community poses several questions for ecologists. overlapping in their resources, such as the grazing High levels of species association occur with high herds in Africa which eat common and widely species packing, as is seen within the Selous game dispersed foods. Sinclair (Sinclair, 1979 as cited in reserve of Tanzania in east Africa. Sinclair (1985) Sinclair, 1985) has found that this seemingly notes that mixed herds are frequently seen in east extensive overlap among these herds have also Africa and Connor and Simberloff (1979) have undergone niche separation. -

Monitoring the Recovery of Wildlife in the Parque Nacional Da Gorongosa Through Aerial Surveys 2000
Monitoring the recovery of wildlife in the Parque Nacional da Gorongosa through aerial surveys 2000 - 2012 A preliminary analysis Dr Marc Stalmans July 2012 Summary • A total of 7 aerial wildlife surveys have been undertaken from 2000 to 2012 in the Gorongosa National Park. These have been sampling surveys covering 9 to 22% of the Park. • The data of these 6 helicopter and 1 fixed-wing (2004) survey have not yet been fully analysed. The data were combined in a single data base of 15 083 individual species occurrence records. These data were also incorporated in a Geographic Information System. • The results clearly indicate that there has been since 2000 a significant increase in wildlife numbers for most species. The vast majority of the wildlife is found in the central and southern part of the fertile Rift Valley. However, densities are much lower in the infertile miombo in the east and west as well as in the Rift Valley closer to human habitation. Where higher levels of protection from illegal hunting are maintained, such as in the Sanctuario, higher densities of wildlife are now recorded even though the recovery started from similar low levels as in other parts of the Park. • The current sampling design has significant shortcomings that make the estimate of overall population numbers problematic, especially for species with relatively low numbers and a clumped distribution (e.g. buffalo and elephant). This makes it difficult to evaluate the current populations against historical wildlife numbers. • The current design is not considered good enough for the quality of data required for ecological research and for the auditing of management performance. -

The Grand Slam
SOUTH AFRICA – PLAINSGAME PACKAGES 2019 (ALL TROPHY FEES IN PACKAGES ARE REFUNDABLE IF NOT TAKEN) Ultimate Hunters The Grand Slam 7 Days + 8 Trophies, all inclusive Wildebeest Slam Impala Slam 1 x Kudu 1 x Gemsbuck 7 Days, 4 trophies, all inclusive 7 Days, 4 trophies, all inclusive 1 x Blue wildebeest 1 x Zebra 1 x Blue Wildebeest 1 x Blesbuck 1 x Impala 1 x Impala 1 x Black Wildebeest 1 x Warthog 1 x Steenbuck or duiker 1 x Black Impala 1 x Golden wildebeest Price: $8 205 1 x Saddleback Impala 1 x King Wildebeest 1 x White Flank Impala Starter Price: $13 800 Price: $9 600 7 Days + 4 Trophies, all inclusive The Daily Rater 1 x Gemsbuck 1 x Blue wildebeest OPTION 1 1 x Impala 1 x Warthog First Timer to Africa 7 Days for $2 240 (hunter) Price: $4 190 7 Days + 5 Trophies, OPTION 2 7 Days for $1 050 (observer) all inclusive OPTION 3 1 x Kudu 7 Days for $525 (children under 12) Lucky Packet 1 x Gemsbuck 7 Days + 5 Trophies 1 x Blue wildebeest Choose 1: Kudu, eland, waterbuck, nyala 1 x Impala Father and Son Choose 1: Gemsbuck, 1 x Warthog 7 Days + 8 Trophies blue wildebeest, zebra, Price: $6 190 Choose 1: Kudu, eland, waterbuck, hartebeest nyala Choose 2: Gemsbuck, blue Choose 1: Blesbuck, Impala Super 6 wildebeest, zebra, hartebeest Choose 1: Warthog, 7 Days + 6 Trophies, 1 x Blesbuck 2 x Impala steenbuck, duiker all inclusive 2 x Warthog Choose 1: Baboon, 1 x Kudu Price: $8 580 monkey, genet, porcupine 1 x Gemsbuck Price: $5 840 Couples 1 x Zebra 7 Days + 6 Trophies, all inclusive 1 x Blesbuck Choose 1: Gemsbuck, blue 1 x Impala wildebeest,