U.O.V.A. BIBLIOTEEK
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Dams in South Africa.Indd
DamsDams inin SouthSouth AfricaAfrica n South Africa we depend mostly on rivers, dams and underground water for our water supply. The country does not get a lot of rain, less than 500 mm a year. In fact, South Africa is one of the 30 driest countries in the world. To make Isure that we have enough water to drink, to grow food and for industries, the government builds dams to store water. A typical dam is a wall of solid material (like concrete, earth and rocks) built across a river to block the flow of the river. In times of excess flow water is stored behind the dam wall in what is known as a reservoir. These dams make sure that communities don’t run out of water in times of drought. About half of South Africa’s annual rainfall is stored in dams. Dams can also prevent flooding when there is an overabundance of water. We have more than 500 government dams in South Africa, with a total capacity of 37 000 million cubic metres (m3) – that’s the same as about 15 million Olympic-sized swimming pools! There are different types of dams: Arch dam: The curved shape of these dams holds back the water in the reservoir. Buttress dam: These dams can be flat or curved, but they always have a series of supports of buttresses on the downstream side to brace the dam. Embankment dam: Massive dams made of earth and rock. They rely on their weight to resist the force of the water. Gravity dam: Massive dams that resist the thrust of the water entirely by their own weight. -

Kai ! Garib Final IDP 2020 2021
KAI !GARIB MUNICIPALITY Integrated Development Plan 2020/2021 “Creating an economically viable and fully developed municipality, which enhances the standard of living of all the inhabitants / community of Kai !Garib through good governance, excellent service delivery and sustainable development.” June 2020 TABLE OF CONTENTS FOREWORD.................................................................................................................1 2. IDP PLANNING PROCESS:......................................................................................2 2.1 IDP Steering Committee:...........................................................................................3 2.2 IDP Representative Forum.........................................................................................3 2.3 Process Overview: Steps & Events:.............................................................................4 2.4 Legislative Framework:…………………………………………………………………………………………...6 3. THE ORGANISATION:............................................................................................15 3.1 Institutional Development………………………………………………………………………………..... 15 3.2 The Vision & Mission:...............................................................................................16 3.3 The Values of Kai !Garib Municipality which guides daily conduct ...............................16 3.4 The functioning of the municipality............................................................................16 3.4.1 Council and council committees..............................................................................16 -

Scientists Brave SA's Mightiest River to Kayak from Source To
Aquatic ecosystems The Orange River forms a green artery of life through the harsh and arid desert along the border of South Africa and Namibia. Courtesy Senqu2SeaCourtesy team Scientists brave SA’s mightiest river to kayak from source to sea When Irrigation Department Director, hile not as substantial to undertake rare extensive field Dr Alfred Dale Lewis, explored the lower as its cousin, the research. “The Orange is the iconic Zambezi, to the north, South African river – long, ancient reaches of the Orange River in December SouthW Africa’s largest river has and traversing varied and incredibly 1913 he walked most of the 400 km-long always captured the imagination of beautiful scenery, from grass moun- journey in one of the hottest years on those who gazed upon it. Local Khoi tain highlands to rocky desert. We named it the Gariep, meaning ‘big wanted to spend an extended period record. Now nearly a century later, three water’ or ‘great river’, while the San’s in nature, experiencing a long rather young researchers of the University of name for it meant ‘Dragon River’. It than a technically difficult adven- Cape Town (UCT) have completed a similar was European commander, Colonel ture,” explains the team. Robert Gordon, who gave the river adventure, traversing South Africa’s its ‘royal’ name, naming the river VALUABLE RESEARCH mightiest river in kayaks from its source after Dutch ruler, Prince William of in the Lesotho mountains to its mouth on Orange, 300 years ago. hile enjoying the scenery For Masters graduate Sam Jack, Wthe team also took time to the West Coast of South Africa. -

Review of Existing Infrastructure in the Orange River Catchment
Study Name: Orange River Integrated Water Resources Management Plan Report Title: Review of Existing Infrastructure in the Orange River Catchment Submitted By: WRP Consulting Engineers, Jeffares and Green, Sechaba Consulting, WCE Pty Ltd, Water Surveys Botswana (Pty) Ltd Authors: A Jeleni, H Mare Date of Issue: November 2007 Distribution: Botswana: DWA: 2 copies (Katai, Setloboko) Lesotho: Commissioner of Water: 2 copies (Ramosoeu, Nthathakane) Namibia: MAWRD: 2 copies (Amakali) South Africa: DWAF: 2 copies (Pyke, van Niekerk) GTZ: 2 copies (Vogel, Mpho) Reports: Review of Existing Infrastructure in the Orange River Catchment Review of Surface Hydrology in the Orange River Catchment Flood Management Evaluation of the Orange River Review of Groundwater Resources in the Orange River Catchment Environmental Considerations Pertaining to the Orange River Summary of Water Requirements from the Orange River Water Quality in the Orange River Demographic and Economic Activity in the four Orange Basin States Current Analytical Methods and Technical Capacity of the four Orange Basin States Institutional Structures in the four Orange Basin States Legislation and Legal Issues Surrounding the Orange River Catchment Summary Report TABLE OF CONTENTS 1 INTRODUCTION ..................................................................................................................... 6 1.1 General ......................................................................................................................... 6 1.2 Objective of the study ................................................................................................ -

South Africa)
FREE STATE PROFILE (South Africa) Lochner Marais University of the Free State Bloemfontein, SA OECD Roundtable on Higher Education in Regional and City Development, 16 September 2010 [email protected] 1 Map 4.7: Areas with development potential in the Free State, 2006 Mining SASOLBURG Location PARYS DENEYSVILLE ORANJEVILLE VREDEFORT VILLIERS FREE STATE PROVINCIAL GOVERNMENT VILJOENSKROON KOPPIES CORNELIA HEILBRON FRANKFORT BOTHAVILLE Legend VREDE Towns EDENVILLE TWEELING Limited Combined Potential KROONSTAD Int PETRUS STEYN MEMEL ALLANRIDGE REITZ Below Average Combined Potential HOOPSTAD WESSELSBRON WARDEN ODENDAALSRUS Agric LINDLEY STEYNSRUST Above Average Combined Potential WELKOM HENNENMAN ARLINGTON VENTERSBURG HERTZOGVILLE VIRGINIA High Combined Potential BETHLEHEM Local municipality BULTFONTEIN HARRISMITH THEUNISSEN PAUL ROUX KESTELL SENEKAL PovertyLimited Combined Potential WINBURG ROSENDAL CLARENS PHUTHADITJHABA BOSHOF Below Average Combined Potential FOURIESBURG DEALESVILLE BRANDFORT MARQUARD nodeAbove Average Combined Potential SOUTPAN VERKEERDEVLEI FICKSBURG High Combined Potential CLOCOLAN EXCELSIOR JACOBSDAL PETRUSBURG BLOEMFONTEIN THABA NCHU LADYBRAND LOCALITY PLAN TWEESPRUIT Economic BOTSHABELO THABA PATSHOA KOFFIEFONTEIN OPPERMANSDORP Power HOBHOUSE DEWETSDORP REDDERSBURG EDENBURG WEPENER LUCKHOFF FAURESMITH houses JAGERSFONTEIN VAN STADENSRUST TROMPSBURG SMITHFIELD DEPARTMENT LOCAL GOVERNMENT & HOUSING PHILIPPOLIS SPRINGFONTEIN Arid SPATIAL PLANNING DIRECTORATE ZASTRON SPATIAL INFORMATION SERVICES ROUXVILLE BETHULIE -

South Africa's Hydropower Options
SUSTAINABLE ENERGY South Africa’s hydropower options by Wilhelm Karanitsch, Andritz Hydro South Africa is still facing the problem of insufficient power generation capacity and a reserve margin which is below international accepted levels. The government is in the approval process for the new IRP 2010 which is considering renewable energy technologies as clean sources of energy that have a significantly lower environmental impact than conventional energy technologies. The IRP 2010 is indicating a greater role in the energy for renewable energy technologies. The National Energy Regulator of South of the energy available from water to be and scattered groups of consumers by Africa has issued Regulatory Guidelines for converted into electricity [1]. a central power station. In areas with a Renewable Energy Feed-in Tariffs ( REFIT). suitable water potential the construction Depending on the head, small/mini hydro Landfill gas power plants, small hydro of a small hydropower plant in island power plants can be classified in the power plants (less than 10 MW), Wind power operation cannot only help to provide following categories: plants and concentrating solar power electricity but also to create jobs. plants qualify as renewable energy power Low head: 2 – 30 m Local industry participation generator. The REFIT should encourage Medium head: 30 – 100 m investment in these technologies. High head: over 100 m For a small hydro power plant the largest South Africa is a dry country which is a cost blocks are the civil works and Kaplan (Propeller, Bulb), Francis, Pelton, turbine/generator set. Costs for a penstock limiting factor for the use of hydropower. -
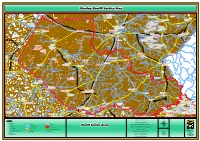
20201101-Fs-Advert Xhariep Sheriff Service Area.Pdf
XXhhaarriieepp SShheerriiffff SSeerrvviiccee AArreeaa UITKYK GRASRANDT KLEIN KAREE PAN VAAL PAN BULTFONTEIN OLIFANTSRUG SOLHEIM WELVERDIEND EDEN KADES PLATKOP ZWAAIHOEK MIDDEL BULT Soutpan AH VLAKPAN MOOIVLEI LOUISTHAL GELUKKIG DANIELSRUST DELFT MARTHINUSPAN HERMANUS THE CRISIS BELLEVUE GOEWERNEURSKOP ROOIPAN De Beers Mine EDEN FOURIESMEER DE HOOP SHEILA KLEINFONTEIN MEGETZANE FLORA MILAMBI WELTEVREDE DE RUST KENSINGTON MARA LANGKUIL ROSMEAD KALKFONTEIN OOST FONTAINE BLEAU MARTINA DORASDEEL BERDINA PANORAMA YVONNE THE MONASTERY JOHN'S LOCKS VERDRIET SPIJT FONTEIN Kimberley SP ROOIFONTEIN OLIFANTSDAM HELPMEKAAR MIMOSA DEALESRUST WOLFPAN ZWARTLAAGTE MORNING STAR PLOOYSBURG BRAKDAM VAALPAN INHOEK CHOE RIETPAN Soetdoring R30 MARIA ATHELOON WATERVAL RUSOORD R709 LOUISLOOTE LAURA DE BAD STOFPUT OPSTAL HERMITAGE WOLVENFONTEIN SUNNYSIDE EERLIJK DORISVILLE ST ZUUR FONTEIN Verkeerdevlei ST LYONSREST R708 UITVAL SANCTUARY SUSANNA BOTHASDAM MERIBA AURORA KALKWAL ^!. VERKEERDEVLEI WATERVAL ZETLAND BELMONT ST SAPS SPITS KOP DIDIMALA LEMOENHOEK WATERVAL ORANGIA SCHOONVLAKTE DWAALHOEK WELTEVREDE GERTJE PAARDEBERG KOPPIES' N8 SANDDAM ZAMENKOMST R64 Nature DIEPHOEK FARMS KARREE KLIMOP MELKVLEY OMDRAAI Mantsopa NU ELYSIUM UMPUKANE HORATIO EUREKA ROODE PAN LK KAMEELPAN KOEDOE`S RAND KLIPFONTEIN DUIKERSDRAAI VLAKLAAGTE ST MIMOSA FAIRFIELD VALAF BEGINSEL Verkeerdevlei SP KOPPIESDAM MELIEFE ZAAIPLAATS PAARDEBERG KARREE DAM ARBEIDSGENOT DOORNLAAGTE EUREKA GELYK TAFELKOP KAREEKOP BOESMANSKOP AHLEN BLAUWKRANS VAN LOVEDALE ALETTA ROODE ESKOL "A" Tokologo NU AANKOMST -

Orange River Project
ORANGE RIVER PROJECT: OvERVIEW South Africa NAMIBIA BOTSWANA Orange-Senqu River Basin Vanderkloof Dam LESOTHO Gariep Dam LOCATION SOUTH AFRICA The Orange River Project (ORP) is the largest scheme in the Orange–Senqu River basin, and includes the two largest dams in South Africa, the Gariep and Vanderkloof. They regulate flows to the Orange River and increase assurance of supply. DESCRIPTION Gariep and Vanderkloof dams were constructed as part of the project, and have a combined storage of 8,500 million m3. The ORP includes several sub-systems. The Orange–Riet Water Scheme.* The Orange–Fish Transfer Tunnel.* The Orange–Vaal Transfer Scheme.* Bloem Water: Pipeline network between Gariep Dam and the towns of Trompsburg, Springfontein and Philippolis. Irrigation abstractions: Between Gariep Dam and downstream of Vanderkloof Dam, up to the confluence of the Vaal and Orange rivers (near the town of Marksdrift). Urban and industrial abstractions: Between Gariep Dam and Marksdrift (including Hopetown and Vanderkloof towns). Support to the Lower Orange Water Management Area Schemes: * Support to most of the demands in the Gariep Dam (© Hendrik van den Berg/Panoramio.com) Lower Orange, including irrigation, urban use and power generation. Caledon–Bloemfontein Government Water Scheme.* * Further details are given on separate pages PURPOSE The purpose of this very complex scheme is to supply demands within several sub-systems, including the Upper and Lower Orange water management areas all the way down to the Orange River mouth, and the Eastern Cape Province. These demands include irrigation, urban, industrial and environmental water requirements. Power generation is also part of the system, including at Gariep and Vanderkloof dams, which contributes to the Eskom national power grid. -

Eskom Audited Annual Results Presentation for the Year Ended 31 March 2011
Eskom Audited Annual Results Presentation for the year ended 31 March 2011 Select Committee on Labour and Public Enterprises 9 May 2012 Disclaimer This presentation does not constitute or form part of and should not be construed as, an offer to sell, or the solicitation or invitation of any offer to buy or subscribe for or underwrite or otherwise acquire, securities of Eskom Holdings Limited (“Eskom”), any holding company or any of its subsidiaries in any jurisdiction or any other person, nor an inducement to enter into any investment activity. No part of this presentation, nor the fact of its distribution, should form the basis of, or be relied on in connection with, any contract or commitment or investment decision whatsoever. This presentation does not constitute a recommendation regarding any securities of Eskom or any other person. Certain statements in this presentation regarding Eskom’s business operations may constitute “forward looking statements.” All statements other than statements of historical fact included in this presentation, including, without limitation, those regarding the financial position, business strategy, management plans and objectives for future operations of Eskom are forward looking statements. Forward-looking statements are not intended to be a guarantee of future results, but instead constitute Eskom’s current expectations based on reasonable assumptions. Forecasted financial information is based on certain material assumptions. These assumptions include, but are not limited to continued normal levels of operating performance and electricity demand in the Distribution and Transmission divisions and operational performance in the Generation and Primary Energy divisions consistent with historical levels, and incremental capacity additions through our Group Capital division at investment levels and rates of return consistent with prior experience, as well as achievements of planned productivity improvements throughout our business activities. -

Media Statement
MEDIA STATEMENT Water on the Rise in Northern Cape due to Heavy Rainfall 02 February 2021 These are the latest dam levels and status of water reservoirs in the Northern Cape according to the Department of Water and Sanitation’s latest water report released as at 01 February 2021. The iconic Vaal Dam which borders Free State and Gauteng, has risen to a storage capacity of 78.36% full with an inflow of 498m3/s and a release of 17.6 m3/s. Bloemhof Dam is at 102% of its storage capacity with a maintained outflow of 101m3/s. The Gariep Dam is 119.7% full with a combined outflow of 2660 m3/s while the Vanderkloof Dam which borders the Free State and Northern Cape provinces is spilling at a capacity of 111.07%, with an inflow of 2660 m3/s and a combined outflow of 1794 m3/s. Due to more rains upstream, the Vaal and Orange Rivers will definitely impact water users downstream of the two river systems. The Douglas Storage Weir has peaked to a high of 136.4% yesterday and spilling at 621 m3/s All water users including irrigation and livestock farmers, fishing, mining and recreation activities along the Orange and Vaal River Systems are advised to stay clear of the possibly flooding rivers. All are urged to remove livestock, water pipes and other working equipment out of the water to avoid damage to property. Communities have also been warned to be on high alert as roads, bridges, dams and water canals in some parts of the province and country are flooded and should be avoided. -

Maximising Hydro-Power Generation Within a Multi-User Water Supply System
226 Hydrocomplexity: New Tools for Solving Wicked Water Problems Kovacs Colloquium, July 2010 (IAHS Publ. 338, 2010) Maximising hydro-power generation within a multi-user water supply system L. BAPELA1, B. MWAKA1, R. CAI1 & H. G. MARÉ2 1 Department of Water Affairs and Forestry, South Africa [email protected] 2 WRP Consulting Engineers (Pty) Ltd, South Africa INTRODUCTION In order to optimise the generation of hydro-power from Gariep and Vanderkloof dams, and to supply the other water users at the required risk levels, a system utilisation agreement was formed between the Department of Water Affairs (DWA) of the Republic of South Africa (RSA) and Eskom (the national energy supplier within the RSA). As part of the agreement between DWA and Eskom, DWA commissioned the Orange River System Annual Operating Analysis study to ensure that all the users are supplied at the required assurance and to determine the volume of water available for the generation of hydro power on an annual basis. Background Gariep and Vanderkloof dams are the two largest dams in the RSA, located in the Orange River, with gross storages of 5343 million m3 and 3187 million m3, respectively. Several water supply sub-systems, including the well known Lesotho Highlands Water Project (LHWP), are located upstream of Gariep and Vanderkloof dams. These sub-systems, some with large transfers, all have an effect on the water availability in Gariep and Vanderkloof dams, also referred to the as the Orange River Project (ORP). The total 2008 demand imposed on the ORP is in the order of 3200 million m3/year-1 and is just lower than the historic firm yield of 3 349 million m3/year-1. -

Hydro Energy Hydroelectric Power Hydro Energy
Hydro Energy Hydroelectric Power Hydro Energy • Hydroelectric power refers to the generation of electric power through the extraction of energy from moving water streams. • A large fraction of radiation reaching the Earth’s surface is absorbed by the oceans, warming them and adding water vapour to the air. Source: Unsplash.com • The water vapour condenses as rain to feed rivers in which dams can be built and hydroelectric turbines installed to extract the energy of the flowing water. • Moving water can be extremely powerful. • The kinetic energy of flowing water can be used to drive hydroelectric turbines to produce electricity. Source: pixabay.com 2 Hydroelectric Power Hydroelectric Power: Hydro Energy Converted To Electricity • Gravity makes water flow from a high to a low place. The moving water contains kinetic energy. Hydroelectric Dam • Hydroelectric power stations are able to transform the kinetic energy in moving water to electrical energy. • In a hydroelectric power station, part of a river’s flow is sent through pipes. • The water then turns the turbines, and the turbines turn the electricity generators. • The water is returned to the river further downstream. • In the conventional system, water is stored behind a dam wall. • The power station is normally situated close to the dam wall. • The water is released on demand, powering huge turbines that generate electricity. 3 Hydroelectric Power Hydroelectric Power in SA • Eskom operates hydroelectric power stations at both the Gariep Dam and the Vanderkloof Dam. • In South Africa, the most important role of these power stations is the storage of ‘electricity’ in case of unexpected demand, or in case of sudden operational disturbances at one of the power stations supplying the regular demand.