A Test of Revisionist Narratives of History in Pre- Famine Ireland
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Download the Potential of Indigenous Wild Foods
The Potential of Indigenous Wild Foods Workshop Proceedings, 22-26 January 2001 April 2001 Funding provided by: USAID/OFDA Implementation provided by: CRS/Southern Sudan Proceeding compilation and editing by: Catherine Kenyatta and Amiee Henderson, USAID contractors The Potential of Indigenous Wild Foods Workshop Proceedings, 22–26 January 2001 April 2001 Funding provided by: USAID/OFDA Implementation provided by: CRS/Southern Sudan Proceeding compilation and editing by: Catherine Kenyatta ([email protected]) and Amiee Henderson ([email protected]), USAID contractors ii Contents Setting the Stage: Southern Sudan Conflict and Transition v Acronyms and Abbreviations ix DAY TWO: JANUARY 23, 2001 Session One Chair: Brian D’Silva, USAID 1 Official Welcome Dirk Dijkerman, USAID/REDSO 1 Overview of the Workshop Brian D’Silva 1 Potential of Indigenous Food Plants to Support and Strengthen Livelihoods in Southern Sudan, Birgitta Grosskinsky, CRS/Sudan, and Caroline Gullick, University College London 2 Discussions/comments from the floor 5 Food Security and the Role of Indigenous Wild Food Plants in South Sudan Mary Abiong Nyok, World Food Programme, Christine Foustino, Yambio County Development Committee, Luka Biong Deng, Sudan Relief and Rehabilitation Association, and Jaden Tongun Emilio, Secretariat of Agriculture and Animal Resources 6 Discussion/comment from the floor 9 Session Two Chair: Brian D’Silva 10 The Wild Foods Database for South Sudan Birgitta Grosskinsky and Caroline Gullick 10 Discussions/Comments from the floor 10 Food -

Hydrogeology of the Burren and Gort Lowlands
KARST HYDROGEOLOGY OF THE BURREN UPLANDS / GORT LOWLANDS Field Guide International Association of Hydrogeologists (IAH) Irish Group 2019 Cover page: View north across Corkscrew Hill, between Lisdoonvarna and Ballyvaughan, one of the iconic Burren vistas. Contributors and Excursion Leaders. Colin Bunce Burren and Cliffs of Moher UNESCO Global Geopark David Drew Department of Geography, Trinity College, Dublin Léa Duran Department of Civil, Structural & Environmental Engineering, Trinity College, Dublin Laurence Gill Department of Civil, Structural & Environmental Engineering, Trinity College, Dublin Bruce Misstear Department of Civil, Structural & Environmental Engineering, Trinity College, Dublin John Paul Moore Fault Analysis Group, Department of Geology, University College Dublin and iCRAG Patrick Morrissey Department of Civil, Structural & Environmental Engineering, Trinity College, Dublin and Roughan O‘Donovan Consulting Engineers David O’Connell Department of Civil, Structural & Environmental Engineering, Trinity College, Dublin Philip Schuler Department of Civil, Structural & Environmental Engineering, Trinity College, Dublin Luka Vucinic Department of Civil, Structural & Environmental Engineering, Trinity College, Dublin and iCRAG Programme th Saturday 19 October 10.00 Doolin Walk north of Doolin, taking in the coast, as well as the Aillwee, Balliny and Fahee North Members, wayboards, chert beds, heterogeneity in limestones, joints and veins, inception horizons, and epikarst David Drew and Colin Bunce, with input from John Paul Moore 13.00 Lunch in McDermotts Bar, Doolin 14.20 Murrooghtoohy Veins and calcite, relationship to caves, groundwater flow and topography John Paul Moore 15.45 Gleninsheen and Poll Insheen Holy Wells, epikarst and hydrochemistry at Poll Insheen Bruce Misstear 17.00 Lisdoonvarna Spa Wells Lisdoonvarna history, the spa wells themselves, well geology, hydrogeology and hydrochemistry, some mysterious heat .. -

1 De Bello Becco
De Bello Becco: a French foundation in the Boyne Valley Geraldine Stout Introduction This paper discusses the history of the little known French Cistercian foundation of de Bello Becco near Drogheda, and the role it played in the commercial development of the Boyne valley and in the international maritime trade during the medieval period. The paper also presents the results of a recent geophysical survey and sets out a course for future investigation of the site. Historical background The Cistercian foundation of de Bello Becco, also known as Beaubec, a daughter house of the convent of de Bello Becco in Normandy, France, was located in the townland of Bey More, Co. Meath (fig. 1, ITM 709546 772877) (St. John Brooks, 1953, 148; McCullen, 2011, 383). There is no known foundation date for this monastic site but a grant by Walter De Lacy to the church of SS Mary and Laurence of Beaubec and the monks residing there on the lands in Gillekeran has been dated to after 1215 (Dryburgh and Smith, 2006, 23). This includes a further grant of a burgage in Mornington (villa Marenari) along with the liberty of a boat for their own use, free from tolls, customs and demands and freedom to access Walter De Lacy’s lands to buy and sell merchandise. The placename Gillekeran suggests that this Cistercian foundation was on an earlier ecclesiastical site possibly associated with St Ciaran (www.monasticon.celt.dias.ie). The 1215 grant is alluded to in a confirmation grant by Henry III in 1235 (Sweetman, 1875, 341). Walter De Lacy bestowed this relatively small grant of land on the Cistercians for the good of his soul, and those of their relatives and ancestors. -

The Politics of Information in Famine Early Warning A
UNIVERSITY OF CALIFORNIA, SAN DIEGO Fixing Famine: The Politics of Information in Famine Early Warning A Dissertation submitted in partial satisfaction of the Requirements for the degree Doctor of Philosophy in Communication by Suzanne M. M. Burg Committee in Charge: Professor Robert B. Horwitz, Chair Professor Geoffrey C. Bowker Professor Ivan Evans Professor Gary Fields Professor Martha Lampland 2008 Copyright Suzanne M. M. Burg, 2008 All rights reserved. The Dissertation of Suzanne M. M. Burg is approved, and it is acceptable in quality and form for publication on microfilm: _______________________________________________________ _______________________________________________________ _______________________________________________________ _______________________________________________________ _______________________________________________________ Chair University of California, San Diego 2008 iii DEDICATION For my past and my future Richard William Burg (1932-2007) and Emma Lucille Burg iv EPIGRAPH I am hungry, O my mother, I am thirsty, O my sister, Who knows my sufferings, Who knows about them, Except my belt! Amharic song v TABLE OF CONTENTS Signature Page……………………………………………………………………. iii Dedication……………………………………………………………………….. iv Epigraph…………………………………………………………………………. v Table of Contents………………………………………………………………... vi List of Acronyms………………………………………………………………… viii List of Figures……………………………………………………………………. xi List of Tables…………………………………………………………………….. xii Acknowledgments……………………………………………………………….. xiii Vita………………………………………………………………………………. -

Your Wedding Day at Buchan Braes Hotel
Your Wedding Day at Buchan Braes Hotel On behalf of all the staff we would like to congratulate you on your upcoming wedding. Set in the former RAF camp, in the village of Boddam, the building has been totally transformed throughout into a contemporary stylish hotel featuring décor and furnishings. The Ballroom has direct access to the landscaped garden which overlooks Stirling Hill, making Buchan Braes Hotel the ideal venue for a romantic wedding. Our Wedding Team is at your disposal to offer advice on every aspect of your day. A wedding is unique and a special occasion for everyone involved. We take pride in individually tailoring all your wedding arrangements to fulfill your dreams. From the ceremony to the wedding reception, our professional staff take great pride and satisfaction in helping you make your wedding day very special. Buchan Braes has 44 Executive Bedrooms and 3 Suites. Each hotel room has been decorated with luxury and comfort in mind and includes all the modern facilities and luxury expected of a 4 star hotel. Your guests can be accommodated at specially reduced rates, should they wish to stay overnight. Our Wedding Team will be delighted to discuss the preferential rates applicable to your wedding in more detail. In order to appreciate what Buchan Braes Hotel has to offer, we would like to invite you to visit the hotel and experience firsthand the four star facilities. We would be delighted to make an appointment at a time suitable to yourself to show you around and discuss your requirements in more detail. -

Establishment of Groundwater Source Protection Zones Martinstown
Establishment of Groundwater Source Protection Zones Martinstown, Ballinvreena Water Supply Scheme December 2010 Prepared by: OCM With contributions from: Dr. Robert Meehan, Ms. Jenny Deakin, Mr. David Ball And with assistance from: Limerick County Council l v Environmental Protection Agency Martinstown Ballinvreena Groundwater SPZ Project description Since the 1980’s, the Geological Survey of Ireland (GSI) has undertaken a considerable amount of work developing Groundwater Protection Schemes throughout the country. Groundwater Source Protection Zones are the surface and subsurface areas surrounding a groundwater source, i.e. a well, wellfield or spring, in which water and contaminants may enter groundwater and move towards the source. Knowledge of where the water is coming from is critical when trying to interpret water quality data at the groundwater source. The Source Protection Zone also provides an area in which to focus further investigation and is an area where protective measures can be introduced to maintain or improve the quality of groundwater. The project “Establishment of Groundwater Source Protection Zones”, led by the Environmental Protection Agency (EPA), represents a continuation of the GSI’s work. A CDM/TOBIN/OCM project team has been retained by the EPA to establish Groundwater Source Protection Zones at monitoring points in the EPA’s National Groundwater Quality Network. A suite of maps and digital GIS layers accompany this report and the reports and maps are hosted on the EPA and GSI websites (www.epa.ie; www.gsi.ie). i Environmental Protection Agency Martinstown Ballinvreena Groundwater SPZ TABLE OF CONTENTS 1 Introduction .............................................................................................................. 1 2 Methodology ............................................................................................................. 1 3 Location, Site Description and Well Head Protection .......................................... -

Irish Freedom Fighters Had Loyal Allies Across the Atlantic Our New Book Commemorates the Role of Connecticut in Ireland’S Long Struggle for Independence
Vol. XXVIII — No. 2 — 2016 Irish freedom fighters had loyal allies across the Atlantic Our new book commemorates the role of Connecticut in Ireland’s long struggle for independence s a contribution to the 2016 centennial celebration of the A Easter Rising in Dublin, the CTIAHS decided to attempt to tell the story of Connecticut links to more than a century of risings and rebellions in Ireland. The end product is a 233-page book — ‘From a land beyond the wave,’ Connecticut’s Irish Rebels, 1798- 1916.’ Shown at right is a black and white picture of the book’s cover which in reality is in full color. The authors are Patrick J. Mahoney, a PhD student at Drew University in New Jersey and a regular columnist for the West Cork People, and Neil Hogan, editor of our newsletter, The Shanachie. The book contains more than 50 articles about the involvement of individual Connecticut Irish men and women in the long strug- gle to win Ireland’s independence. Their involvement included everything from writing poetry to delivering orations to raising funds, rescuing prisoners, dynamite attacks in England and mili- tary invasions of Canada. Also described in the book are activities of various Irish organi- zations in Connecticut to further the cause of Irish independence including the Emmet Monument Association, Clan na Gael, the An- cient Order of Hibernians and its women’s auxiliary, the American affiliate of the Irish Land League and the Friends of Irish Freedom. The book also tells the story of how numerous Irish nationalist leaders visited Connecticut and other areas of the United States to strengthen the bonds between Irish immigrants and those they left behind. -
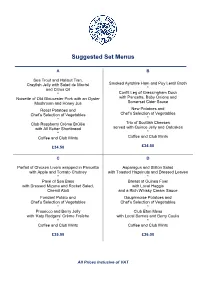
Scottish Menus
Suggested Set Menus A B Sea Trout and Halibut Tian, Crayfish Jelly with Salad de Maché Smoked Ayrshire Ham and Puy Lentil Broth and Citrus Oil * * Confit Leg of Gressingham Duck Noisette of Old Gloucester Pork with an Oyster with Pancetta, Baby Onions and Somerset Cider Sauce Mushroom and Honey Jus Roast Potatoes and New Potatoes and Chef’s Selection of Vegetables Chef’s Selection of Vegetables * * Club Raspberry Crème Brûlée Trio of Scottish Cheeses with All Butter Shortbread served with Quince Jelly and Oatcakes * * Coffee and Club Mints Coffee and Club Mints £34.50 £34.50 C D Parfait of Chicken Livers wrapped in Pancetta Asparagus and Stilton Salad with Apple and Tomato Chutney with Toasted Hazelnuts and Dressed Leaves * * Pavé of Sea Bass Breast of Guinea Fowl with Dressed Mizuna and Rocket Salad, with Local Haggis Chervil Aïoli and a Rich Whisky Cream Sauce Fondant Potato and Dauphinoise Potatoes and Chef’s Selection of Vegetables Chef’s Selection of Vegetables * * Prosecco and Berry Jelly Club Eton Mess with ‘Katy Rodgers’ Crème Fraîche with Local Berries and Berry Coulis * * Coffee and Club Mints Coffee and Club Mints £35.00 £36.00 All Prices Inclusive of VAT Suggested Set Menus E F Confit of Duck, Guinea Fowl and Apricot Rosettes of Loch Fyne Salmon, Terrine, Pea Shoot and Frissée Salad Lilliput Capers, Lemon and Olive Dressing * * Escalope of Seared Veal, Portobello Mushroom Tournedos of Border Beef Fillet, and Sherry Cream with Garden Herbs Fricasée of Woodland Mushrooms and Arran Mustard Château Potatoes and Chef’s Selection -

Heritage Council Annual Report 2014
Annual Report 2014 www.heritagecouncil.ie CONTENTS © The Heritage Council 2015 All rights reserved. No part of this book may be printed or reproduced or utilised in any electronic, mechanical, or other means, now known or heretoafter invented, including photocopying or licence permitting restricted copying in Ireland issued by the Irish Copyright Licencing Agency Ltd., The Writers Centre, 19 Parnell Square, Dublin 1 Published by the Heritage Council CHAIRMAN’S WELCOME 2 The Heritage Council of Ireland Series CHIEF EXECUTIVE’S REPORT 2014 3 ISBN 978-1-906304-30-0 HERITAGE COUNCIL BOARD MEMBERS AND STAFF 2014 5 THE HERITAGE COUNCIL – 2014 IN FIGURES 6 The Heritage Council is extremely grateful to the following organisations and individuals for supplying additional photographs, images and diagrams used in the Annual Report 2014: 1 BACKGROUND TO HERITAGE & THE HERITAGE COUNCIL – STRATEGIC OBJECTIVES, RESOURCES AND SUSTAINABILITY SUSTAINABILITY 8 Andrew Power (Heritage Week), Birdwatch Ireland, Burrenbeo Trust (Dr. Brendan Dunford), Brady Shipman Martin, Clare Keogh (Cork City Council), Clive Wasson Photography (Donegal), David Jordan (Co. Carlow), Europa Nostra, the Irish Planning Institute (IPI), Kilkenny Tourism, 2 OUR PERFORMANCE IN 2014 – NATIONAL AND INTERNATIONAL AWARDS RECEIVED BY THE HERITAGE COUNCIL 20 Dr. Liam Lysaght (Director, National Biodiversity Data Centre), Michael Martin (Co. Carlow), Michael Scully (Laois), Valerie O’Sullivan Photography (Co. Kerry), Pat Moore (Co. Kilkenny) and The Paul Hogarth Company (Belfast and Dublin). 3 2014 KEY HIGHLIGHTS – POLICY ADVICE, PROGRAMMES AND PROJECTS 24 Images used on pages 20, 30, 64 and 82 © Photographic Unit, National Monuments Service 4 SUPPORTING EMPLOYMENT AND JOB CREATION (HC OBJECTIVE NO. -

8 Solo Championship Results Oireachtas
SOLO CHAMPIONSHIP RESULTS OIREACHTAS RINCE ULADH 2014 Comórtas 8 Minor Mixed Championship 7 - 8 1st 27 Fionn Higgins S.R. Breffni 2nd 30 Chloe Moore S.R. McConomy Bradley 3rd 37 Niamh Murray S.R. McConomy Bradley 4th 36 Carmen McAleer S.R. Clann McCaul 5th 33 Shannon Clawson Ard-Rialla O'Donoghue 6th 28 Lily Blachut Ursula Moore Academy 7th 21 Jemma McNamara S.R. Ni Chearra - O Baolain 8th 32 Naoise McCrory Doherty Petri 9th 29 Mary-Jayne Burns Taylor 10th 23 Alicja Drogez Ursula Moore Academy 11th 34 Katie O'Neill McConomy 12th 35 Seoda Wang McNelis - Cunningham - Boyle 13th 11 Aoife McDermott Flanagan McMahon Moore 14th 2 Katie Scarlett McCaul Flanagan McMahon Moore 15th 4 Roise McCormick Lorraine Scallon 16th 31 Shay Maby Smith McCarron 17th 14 Pearse Rainey Doherty Petri 18th Tie 18 Eabha Wade Domican 18th Tie 24 Natteya Cruickshank S.R. McConomy Bradley SOLO CHAMPIONSHIP RESULTS OIREACHTAS RINCE ULADH 2014 Comórtas 9 Minor Mixed Championship 8 - 9 1st 10 Blanaid O'Neill Doherty Petri 2nd 52 Lara Magee S.R. McConomy Bradley 3rd 29 Enya Higgins S.R. Breffni 4th 14 Óisin Lennon S.R. Mona Ni Rodaigh 5th 47 Amber-Dee Leonard Hanly Ard-Rialla O'Donoghue 6th 45 Hannah Clarke S.R. Realta 7th 41 Renee Leonard Crotty Ard-Rialla O'Donoghue 8th 26 Daimhín Pritchard McConomy 9th 33 Grace Kelly S.R. Ni Chearra - O Baolain 10th 40 Rebecca Whyte Ursula Moore Academy 11th Tie 37 Leona Mulvey Carol Leavy 11th Tie 38 Paige Lockhart Taylor 13th 36 Neasa Boyle McNelis - Cunningham - Boyle 14th 50 Jude McGibbon Doherty Petri 15th 31 Caoimhe Gallagher S.R. -

Chapter 7: Infrastructure
Navan Development Plan 2009-2015 November 2009 CHAPTER 7: INFRASTRUCTURE SECTIONS IN THIS CHAPTER 7.1 Transportation-Introduction 7.2 Goals 7.3 Roads 7.4 Pedestrians and Cyclists 7.5 Public Transport 7.6 Park & Ride, Parking, Loading & Taxi Ranks 7.7 Street Furniture & Lighting 7.8 Keeping the Street Clean 7.9 Water Supply & Sewerage Treatment & Disposal 7.10 Flooding 7.11 Sustainable Urban Drainage Systems 7.12 Telecommunications 7.13 Broadband 7.14 Solid Waste Collection & Disposal 7.15 Energy 161 Navan Development Plan 2009-2015 November 2009 7.1 Transportation-Introduction The context for this chapter is provided in Chapter Two of this Development Plan. The key to Navan’s sustainable growth is the development of an integrated land use and transportation system which provides for a development pattern that protects Navan’s architectural and natural heritage while providing for growth and economic development. The integration of land use and transportation policies, both in the town centre and outer suburban areas underpins the transportation strategy as outlined below. 7.2 Goals It is the goal of Navan Town Council and Meath County Council to create vibrant urban areas with good provision of public transport, reduced traffic congestion and an attractive urban centre, which is not dominated by the car. It is the main objective of the Planning Authorities to create an integrated and environmentally sound system, with particular emphasis on: Accessibility and choice of transport for all; Strong integration between transportation planning -

A Dinner at the Governor's Palace, 10 September 1770
W&M ScholarWorks Dissertations, Theses, and Masters Projects Theses, Dissertations, & Master Projects 1998 A Dinner at the Governor's Palace, 10 September 1770 Mollie C. Malone College of William & Mary - Arts & Sciences Follow this and additional works at: https://scholarworks.wm.edu/etd Part of the American Studies Commons Recommended Citation Malone, Mollie C., "A Dinner at the Governor's Palace, 10 September 1770" (1998). Dissertations, Theses, and Masters Projects. Paper 1539626149. https://dx.doi.org/doi:10.21220/s2-0rxz-9w15 This Thesis is brought to you for free and open access by the Theses, Dissertations, & Master Projects at W&M ScholarWorks. It has been accepted for inclusion in Dissertations, Theses, and Masters Projects by an authorized administrator of W&M ScholarWorks. For more information, please contact [email protected]. A DINNER AT THE GOVERNOR'S PALACE, 10 SEPTEMBER 1770 A Thesis Presented to The Faculty of the Department of American Studies The College of William and Mary in Virginia In Partial Fulfillment Of the Requirements for the Degree of Master of Arts by Mollie C. Malone 1998 APPROVAL SHEET This thesis is submitted in partial fulfillment of the requirements for the degree of Master of Arts 'JYIQMajl C ^STIclU ilx^ Mollie Malone Approved, December 1998 P* Ofifr* * Barbara (farson Grey/Gundakerirevn Patricia Gibbs Colonial Williamsburg Foundation TABLE OF CONTENTS Page ACKNOWLEDGEMENTS iv ABSTRACT V INTRODUCTION 2 HISTORIOGRAPHY 5 A DINNER AT THE GOVERNOR’S PALACE, 10 SEPTEMBER 1770 17 CONCLUSION 45 APPENDIX 47 BIBLIOGRAPHY 73 i i i ACKNOWLEDGMENTS I want to thank Professor Barbara Carson, under whose guidance this paper was completed, for her "no-nonsense" style and supportive advising throughout the project.