Weed Management in Conventional, No-Till, and Transgenic Corn With
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

2,4-Dichlorophenoxyacetic Acid
2,4-Dichlorophenoxyacetic acid 2,4-Dichlorophenoxyacetic acid IUPAC (2,4-dichlorophenoxy)acetic acid name 2,4-D Other hedonal names trinoxol Identifiers CAS [94-75-7] number SMILES OC(COC1=CC=C(Cl)C=C1Cl)=O ChemSpider 1441 ID Properties Molecular C H Cl O formula 8 6 2 3 Molar mass 221.04 g mol−1 Appearance white to yellow powder Melting point 140.5 °C (413.5 K) Boiling 160 °C (0.4 mm Hg) point Solubility in 900 mg/L (25 °C) water Related compounds Related 2,4,5-T, Dichlorprop compounds Except where noted otherwise, data are given for materials in their standard state (at 25 °C, 100 kPa) 2,4-Dichlorophenoxyacetic acid (2,4-D) is a common systemic herbicide used in the control of broadleaf weeds. It is the most widely used herbicide in the world, and the third most commonly used in North America.[1] 2,4-D is also an important synthetic auxin, often used in laboratories for plant research and as a supplement in plant cell culture media such as MS medium. History 2,4-D was developed during World War II by a British team at Rothamsted Experimental Station, under the leadership of Judah Hirsch Quastel, aiming to increase crop yields for a nation at war.[citation needed] When it was commercially released in 1946, it became the first successful selective herbicide and allowed for greatly enhanced weed control in wheat, maize (corn), rice, and similar cereal grass crop, because it only kills dicots, leaving behind monocots. Mechanism of herbicide action 2,4-D is a synthetic auxin, which is a class of plant growth regulators. -

Environmental Fate of Imidazolinone Herbicides and Their Enantiomers
Title Environmental Fate of Imidazolinone Herbicides and Their Enantiomers in Soil and Water Mohammadkazem Ramezani B.Sc. Agronomy, M.Sc. Weed Science This thesis is presented for the degree of Doctorate of Philosophy of the University of Adelaide School of Agriculture, Food & Wine The University of Adelaide Waite Campus, South Australia 2007 Declaration This work contains no material which has been accepted for the award of any other degree or diploma in any university or other tertiary institution and, to the best of my knowledge and belief, contains no material previously published or written by another person, except where due reference has been made in the text. I give consent to this copy of my thesis, when deposited in the University Library, being made available for loan and photocopying. Mohammadkazem Ramezani ii Abstract Imidazolinones represent a new class of herbicides with low mammalian toxicity that can be used at low application rates, either pre- or post-emergence for the control of a wide range of weeds in broadleaf and cereal crops, and non-crop situations. All imidazolinone herbicides are chiral, containing two enantiomers that derive from the chiral centre of the imidazolinone ring. The inhibitory activity of the R(+) enantiomer is nearly eight times greater than that of the S(-) enantiomer. The use of imidazolinone herbicides has increased in recent years in Australia owing to increased popularity of pulses and the introduction of imidazolinone-tolerant canola and wheat. Concerns have been raised about the potential carry over damage to the subsequent crops grown in rotation with legumes and herbicide tolerant crops. -

Herbicide Mode of Action Table High Resistance Risk
Herbicide Mode of Action Table High resistance risk Chemical family Active constituent (first registered trade name) GROUP 1 Inhibition of acetyl co-enzyme A carboxylase (ACC’ase inhibitors) clodinafop (Topik®), cyhalofop (Agixa®*, Barnstorm®), diclofop (Cheetah® Gold* Decision®*, Hoegrass®), Aryloxyphenoxy- fenoxaprop (Cheetah®, Gold*, Wildcat®), fluazifop propionates (FOPs) (Fusilade®), haloxyfop (Verdict®), propaquizafop (Shogun®), quizalofop (Targa®) Cyclohexanediones (DIMs) butroxydim (Factor®*), clethodim (Select®), profoxydim (Aura®), sethoxydim (Cheetah® Gold*, Decision®*), tralkoxydim (Achieve®) Phenylpyrazoles (DENs) pinoxaden (Axial®) GROUP 2 Inhibition of acetolactate synthase (ALS inhibitors), acetohydroxyacid synthase (AHAS) Imidazolinones (IMIs) imazamox (Intervix®*, Raptor®), imazapic (Bobcat I-Maxx®*, Flame®, Midas®*, OnDuty®*), imazapyr (Arsenal Xpress®*, Intervix®*, Lightning®*, Midas®* OnDuty®*), imazethapyr (Lightning®*, Spinnaker®) Pyrimidinyl–thio- bispyribac (Nominee®), pyrithiobac (Staple®) benzoates Sulfonylureas (SUs) azimsulfuron (Gulliver®), bensulfuron (Londax®), chlorsulfuron (Glean®), ethoxysulfuron (Hero®), foramsulfuron (Tribute®), halosulfuron (Sempra®), iodosulfuron (Hussar®), mesosulfuron (Atlantis®), metsulfuron (Ally®, Harmony®* M, Stinger®*, Trounce®*, Ultimate Brushweed®* Herbicide), prosulfuron (Casper®*), rimsulfuron (Titus®), sulfometuron (Oust®, Eucmix Pre Plant®*, Trimac Plus®*), sulfosulfuron (Monza®), thifensulfuron (Harmony®* M), triasulfuron (Logran®, Logran® B-Power®*), tribenuron (Express®), -

Manuka Oil As a Potential Natural Herbicide
Manuka Oil as a Potential Natural Herbicide Franck E. Dayan and Daniel K. Owens USDA-ARS Natural Products Utilization Research Unit P.O. Box 8048, University, MS 38677 [email protected] In 1977, Gray observed that bottlebrush plant (Callistemon citrinus) repressed the growth of plants in its surroundings. Crude extracts from this plant caused the bleaching of grass weeds. He identified the active component as leptospermone, a natural triketone structure with no known biological activity that had been reported in a number of Australasian shrubs. Leptospermone was moderately active in greenhouse tests, controlling mostly small-seeded grass weeds. This natural product and a small number of synthetic structural analogs were patented as herbicides in 1980. A few years later, a separate group at the Western Research Center was generating analogs of the cyclohexanedione herbicide sethoxydim, an inhibitor or acetyl- coenzyme-A carboxylase. Some of the second generation herbicidal derivatives with a dimedone backbone caused bleaching symptoms similar to leptospermone. Combination of the syncarpic acid of leptospermone to this chemistry ultimately served as the basis for the development of the triketone synthetic herbicides (Fig. 1). Fig. 1. Struct ure s of the n a tural trik et o n e leptosperm one a nd t w o s y nthetic analogues that are sold as commer ci a l he r bic ides. Natural β-triketones are common in many Australasian woody plants (e.g., Leptospermum, Eucalyptus, Melaceuca, etc...). Steam distilled manuka oil account for 0.3% of the dried weight of L. scoparium. However, the amount of β-triketone present in these oils varies wildly across New Zealand. -

Exposure to Herbicides in House Dust and Risk of Childhood Acute Lymphoblastic Leukemia
Journal of Exposure Science and Environmental Epidemiology (2013) 23, 363–370 & 2013 Nature America, Inc. All rights reserved 1559-0631/13 www.nature.com/jes ORIGINAL ARTICLE Exposure to herbicides in house dust and risk of childhood acute lymphoblastic leukemia Catherine Metayer1, Joanne S. Colt2, Patricia A. Buffler1, Helen D. Reed3, Steve Selvin1, Vonda Crouse4 and Mary H. Ward2 We examine the association between exposure to herbicides and childhood acute lymphoblastic leukemia (ALL). Dust samples were collected from homes of 269 ALL cases and 333 healthy controls (o8 years of age at diagnosis/reference date and residing in same home since diagnosis/reference date) in California, using a high-volume surface sampler or household vacuum bags. Amounts of agricultural or professional herbicides (alachlor, metolachlor, bromoxynil, bromoxynil octanoate, pebulate, butylate, prometryn, simazine, ethalfluralin, and pendimethalin) and residential herbicides (cyanazine, trifluralin, 2-methyl-4- chlorophenoxyacetic acid (MCPA), mecoprop, 2,4-dichlorophenoxyacetic acid (2,4-D), chlorthal, and dicamba) were measured. Odds ratios (OR) and 95% confidence intervals (CI) were estimated by logistic regression. Models included the herbicide of interest, age, sex, race/ethnicity, household income, year and season of dust sampling, neighborhood type, and residence type. The risk of childhood ALL was associated with dust levels of chlorthal; compared to homes with no detections, ORs for the first, second, and third tertiles were 1.49 (95% CI: 0.82–2.72), 1.49 (95% CI: 0.83–2.67), and 1.57 (95% CI: 0.90–2.73), respectively (P-value for linear trend ¼ 0.05). The magnitude of this association appeared to be higher in the presence of alachlor. -

Efficacy of Imazapic/Imazapyr and Other Herbicides in Mixtures for The
Efficacy of imazapic/imazapyr and other herbicides in mixtures for the control of Digitaria insularis prior to soybean sowing Efectividad de imazapic/imazapyr y otros herbicidas en mezclas para el control de Digitaria insularis en pre-siembra de soya Alfredo Junior Paiola Albrecht1, Leandro Paiola Albrecht1, André Felipe Moreira Silva²*, Romulo Augusto Ramos³, Everson Pedro Zeny³, Juliano Bortoluzzi Lorenzetti4, Maikon Tiago Yamada Danilussi4, and Arthur Arrobas Martins Barroso4 ABSTRACT RESUMEN Herbicide mixtures, use of multiple sites of action, and other Las mezclas entre herbicidas, el uso de múltiples sitios de acción weed management practices are necessary to avoid cases of y otras prácticas de manejo de malezas son necesarias para biotype resistance. The aim of this study was to evaluate the evitar otros casos de resistencia de biotipos. El objetivo de este efficiency of imazapic/imazapyr and other herbicides in mix- estudio fue evaluar la eficiencia de imazapic/imazapyr y otros tures to control Digitaria insularis at burndown before soybean herbicidas en mezclas para controlar Digitaria insularis en la sowing. This field research was conducted in Umuarama, State desecación antes de la siembra de soya. Esta investigación de of Parana (PR), Brazil, in the 2018/19 soybean season. The ex- campo se realizó en Umuarama, Estado de Paraná (PR), Brasil, periment was conducted in a randomized block experimental en la cosecha de soya de 2018/19. El experimento se realizó en design with four replicates and 11 treatments composed of the un diseño experimental de bloques al azar, con cuatro repe- application of glyphosate, clethodim, haloxyfop, imazapic/ ticiones y 11 tratamientos, compuestos por la aplicación de imazapyr, glufosinate, 2,4-dichlorophenoxyacetic acid (2,4-D), glifosato, cletodim, haloxifop, imazapic/imazapir, glufosinato, dicamba, triclopyr, and saflufenacil, in mixtures. -

INDEX to PESTICIDE TYPES and FAMILIES and PART 180 TOLERANCE INFORMATION of PESTICIDE CHEMICALS in FOOD and FEED COMMODITIES
US Environmental Protection Agency Office of Pesticide Programs INDEX to PESTICIDE TYPES and FAMILIES and PART 180 TOLERANCE INFORMATION of PESTICIDE CHEMICALS in FOOD and FEED COMMODITIES Note: Pesticide tolerance information is updated in the Code of Federal Regulations on a weekly basis. EPA plans to update these indexes biannually. These indexes are current as of the date indicated in the pdf file. For the latest information on pesticide tolerances, please check the electronic Code of Federal Regulations (eCFR) at http://www.access.gpo.gov/nara/cfr/waisidx_07/40cfrv23_07.html 1 40 CFR Type Family Common name CAS Number PC code 180.163 Acaricide bridged diphenyl Dicofol (1,1-Bis(chlorophenyl)-2,2,2-trichloroethanol) 115-32-2 10501 180.198 Acaricide phosphonate Trichlorfon 52-68-6 57901 180.259 Acaricide sulfite ester Propargite 2312-35-8 97601 180.446 Acaricide tetrazine Clofentezine 74115-24-5 125501 180.448 Acaricide thiazolidine Hexythiazox 78587-05-0 128849 180.517 Acaricide phenylpyrazole Fipronil 120068-37-3 129121 180.566 Acaricide pyrazole Fenpyroximate 134098-61-6 129131 180.572 Acaricide carbazate Bifenazate 149877-41-8 586 180.593 Acaricide unclassified Etoxazole 153233-91-1 107091 180.599 Acaricide unclassified Acequinocyl 57960-19-7 6329 180.341 Acaricide, fungicide dinitrophenol Dinocap (2, 4-Dinitro-6-octylphenyl crotonate and 2,6-dinitro-4- 39300-45-3 36001 octylphenyl crotonate} 180.111 Acaricide, insecticide organophosphorus Malathion 121-75-5 57701 180.182 Acaricide, insecticide cyclodiene Endosulfan 115-29-7 79401 -
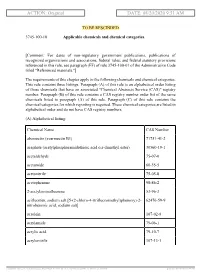
ACTION: Original DATE: 08/20/2020 9:51 AM
ACTION: Original DATE: 08/20/2020 9:51 AM TO BE RESCINDED 3745-100-10 Applicable chemicals and chemical categories. [Comment: For dates of non-regulatory government publications, publications of recognized organizations and associations, federal rules, and federal statutory provisions referenced in this rule, see paragraph (FF) of rule 3745-100-01 of the Administrative Code titled "Referenced materials."] The requirements of this chapter apply to the following chemicals and chemical categories. This rule contains three listings. Paragraph (A) of this rule is an alphabetical order listing of those chemicals that have an associated "Chemical Abstracts Service (CAS)" registry number. Paragraph (B) of this rule contains a CAS registry number order list of the same chemicals listed in paragraph (A) of this rule. Paragraph (C) of this rule contains the chemical categories for which reporting is required. These chemical categories are listed in alphabetical order and do not have CAS registry numbers. (A) Alphabetical listing: Chemical Name CAS Number abamectin (avermectin B1) 71751-41-2 acephate (acetylphosphoramidothioic acid o,s-dimethyl ester) 30560-19-1 acetaldehyde 75-07-0 acetamide 60-35-5 acetonitrile 75-05-8 acetophenone 98-86-2 2-acetylaminofluorene 53-96-3 acifluorfen, sodium salt [5-(2-chloro-4-(trifluoromethyl)phenoxy)-2- 62476-59-9 nitrobenzoic acid, sodium salt] acrolein 107-02-8 acrylamide 79-06-1 acrylic acid 79-10-7 acrylonitrile 107-13-1 [ stylesheet: rule.xsl 2.14, authoring tool: RAS XMetaL R2_0F1, (dv: 0, p: 185720, pa: -

Jamesdanieljonesiiithesis.Doc-After Defense
The Evaluation of HPPD-Inhibitors for Full-Season Control of Morningglory (Ipomoea) Species in Corn (Zea mays L.) by James Daniel Jones III A thesis submitted to the Graduate Faculty of Auburn University in partial fulfillment of the requirements for the Degree of Master of Science Auburn, Alabama December 15, 2018 Keywords: Ipomoea, corn, HPPD, postemergence Approved by Dr. Dennis Delaney, Chair, Extension Specialist, Crop, Soil, and Environmental Sciences Dr. Andrew Price, Affiliate Associate Professor, Crop, Soil, and Environmental Sciences Dr. Audrey Gamble, Assistant Professor and Extension Specialist, Crop, Soil, and Environmental Sciences i Abstract Due to late-season morningglory harvest interference concerns in corn, field studies were conducted in 2017 and 2018 at the Prattville Agricultural Research Unit in Prattville, Alabama and at the Sand Mountain Research and Extension Center in Crossville, Alabama to evaluate late season control of morningglory species using HPPD- inhibitors postemergence (POST) applied alone, following atrazine preemergence (PRE), or in combination with atrazine. Additionally, Amaranthus spp. and Senna spp. were evaluated for control. An incomplete randomized design with a split-plot treatment arrangement with four replications was utilized. The trial was divided into two sections: one with a PRE application of atrazine and a second without a preemergence application of atrazine. Eleven herbicides were applied POST without atrazine including: tembotrione; mesotrione; topramezone+dimethenamid; mesotrione+S- metolachlor+glyphosate; tembotrione+thiencarbazone; topramezone; mesotrione+S- metolachlor+atrazine; mesotrione+S-metolachlor+atrazine+bicyclopyrone; mesotrione+nicosulfuron; isoxaflutole; isoxaflutole+thiencarbazone-methyl; non-treated with atrazine applied PRE, and a true non-treated check. The same herbicides, excluding the two treatments that contain atrazine in the premixture, were also applied with atrazine. -

Comparison of Imazapyr and Imazamox for Control of Parrotfeather (Myriophyllum Aquaticum (Vell.) Verdc.)
J. Aquat. Plant Manage. 45: 132-136 Comparison of Imazapyr and Imazamox for Control of Parrotfeather (Myriophyllum aquaticum (Vell.) Verdc.) RYAN M. WERSAL1 AND JOHN D. MADSEN1,2 INTRODUCTION techniques (Moreira et al. 1999). To date, chemical control has been the most effective method for controlling infesta- Parrotfeather (Myriophyllum aquaticum (Vell. Verdc) is an in- tions of M. aquaticum. Contact herbicides such as diquat vasive aquatic plant to the United States that is native to South (6,7-dihydrodipyrido (1,2-a:2’,1’-c) pyrazinedium dibro- America. Myriophyllum aquaticum is described as “stout, stems mide) and endothall (7-oxabicyclo (2.2.1) heptane-2,3-dicar- moderately elongate, partially submersed but with portions of boxylic acid) have been evaluated with mixed results leafy branches emersed (Godfrey and Wooten 1981). Emer- (Moreira et al. 1999, Westerdahl and Getsinger 1988). Con- gent leaves are whorled, stiff, usually with 20 or more linear fil- tact herbicides are typically effective for short-term control, iform divisions, appearing feather-like and grayish green. but significant regrowth of M. aquaticum typically occurs and Submersed shoots are comprised of whorls of four to six fila- multiple applications are necessary (Moreira et al. 1999). mentous, pectinate, red or orange, leaves arising from each Therefore, the use of a systemic herbicide may be more ef- node. Flowers are all pistillate, borne in the axils of unreduced fective in controlling this species. leaves (Godfrey and Wooten 1981). Myriophyllum aquaticum is Imazapyr (2-[4,5-dihydro-4-methyl-4-(1-methylethyl)5-oxo- dioecious, however only pistillate plants are found outside of 1H-imazol-2-yl]-3-pyridinecarboxylic acid) is a systemic herbi- its native range (Sutton 1985). -

AP-42, CH 9.2.2: Pesticide Application
9.2.2PesticideApplication 9.2.2.1General1-2 Pesticidesaresubstancesormixturesusedtocontrolplantandanimallifeforthepurposesof increasingandimprovingagriculturalproduction,protectingpublichealthfrompest-bornediseaseand discomfort,reducingpropertydamagecausedbypests,andimprovingtheaestheticqualityofoutdoor orindoorsurroundings.Pesticidesareusedwidelyinagriculture,byhomeowners,byindustry,andby governmentagencies.Thelargestusageofchemicalswithpesticidalactivity,byweightof"active ingredient"(AI),isinagriculture.Agriculturalpesticidesareusedforcost-effectivecontrolofweeds, insects,mites,fungi,nematodes,andotherthreatstotheyield,quality,orsafetyoffood.Theannual U.S.usageofpesticideAIs(i.e.,insecticides,herbicides,andfungicides)isover800millionpounds. AiremissionsfrompesticideusearisebecauseofthevolatilenatureofmanyAIs,solvents, andotheradditivesusedinformulations,andofthedustynatureofsomeformulations.Mostmodern pesticidesareorganiccompounds.EmissionscanresultdirectlyduringapplicationorastheAIor solventvolatilizesovertimefromsoilandvegetation.Thisdiscussionwillfocusonemissionfactors forvolatilization.Thereareinsufficientdataavailableonparticulateemissionstopermitemission factordevelopment. 9.2.2.2ProcessDescription3-6 ApplicationMethods- Pesticideapplicationmethodsvaryaccordingtothetargetpestandtothecroporothervalue tobeprotected.Insomecases,thepesticideisapplieddirectlytothepest,andinotherstothehost plant.Instillothers,itisusedonthesoilorinanenclosedairspace.Pesticidemanufacturershave developedvariousformulationsofAIstomeetboththepestcontrolneedsandthepreferred -

Imazapyr Human Health and Ecological Risk Assessment FINAL REPORT
SERA TR-052-29-03a Imazapyr Human Health and Ecological Risk Assessment FINAL REPORT Submitted to: Paul Mistretta, COR USDA/Forest Service, Southern Region 1720 Peachtree RD, NW Atlanta, Georgia 30309 USDA Forest Service Contract: AG-3187-C-06-0010 USDA Forest Order Number: AG-43ZP-D-11-0012 SERA Internal Task No. 52-29 Submitted by: Patrick R. Durkin Syracuse Environmental Research Associates, Inc. 8125 Solomon Seal Manlius, New York 13104 E-Mail: [email protected] Home Page: www.sera-inc.com December 16, 2011 Table of Contents LIST OF FIGURES ...................................................................................................................... vii LIST OF TABLES ........................................................................................................................ vii LIST OF APPENDICES ............................................................................................................... vii ACRONYMS, ABBREVIATIONS, AND SYMBOLS .............................................................. viii COMMON UNIT CONVERSIONS AND ABBREVIATIONS .................................................... x CONVERSION OF SCIENTIFIC NOTATION ........................................................................... xi EXECUTIVE SUMMARY .......................................................................................................... xii 1. INTRODUCTION ...................................................................................................................... 1 1.1. Chemical Specific Information