A Molecular Phylogeny of the Peacock-Pheasants (Galliformes: Polyplectron Spp.) Indicates Loss and Reduction of Ornamental Traits and Display Behaviours
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Congo Peacock – an Update
Congo Peacock – an update The discovery of Congo Peacock Afropavo congensis was one of the most sensational ornithological events of the 20 th century, partly because such a large bird had eluded discovery and partly because of its potential affinity with Asiatic species (1). It was in 1936 that James Chapin found two mounted specimens put away on top of a cabinet at the Belgian Congo Museum (now Royal Museum for Central Africa in Tervuren, near Brussels), having been misidentified as Pavo cristatus for 20 years, and thus considered uninteresting (2). It was rediscovered and photographed by wildlife film-maker Alan Root in 1993 (3, 4). It has remained an elusive and rarely seen and photographed species so it was with great interest and excitement to receive this photograph of a female Congo Peacock located by Mark Van Beirs (http://www.birdquest-tours.com/news ) ABC member Michel Louette, the former head of Ornithology at the RMCA writes to say that his late student Emile Mulotwa at the University of Kisangani, worked on this species for his doctoral thesis in 2008. During this work, he made many sightings (5-7). But Emile died sadly in a plane accident in 2011. References 1. Urban, E.K., Fry, C.H. and Keith, S. (1986) The Birds of Africa Volume II, pp. 11-13. 2. Chapin, J.P. (1936) A new peacock-like bird from the Belgian Congo. Rev. Zool. Bot. Afr. 29, pp. 1-6. 3. Fanshawe, J.H. (1994): Elusive Congo Peacock found by filmmaker Alan Root. ABC Bulletin 1(1) p 27. -

A Molecular Phylogeny of the Pheasants and Partridges Suggests That These Lineages Are Not Monophyletic R
Molecular Phylogenetics and Evolution Vol. 11, No. 1, February, pp. 38–54, 1999 Article ID mpev.1998.0562, available online at http://www.idealibrary.com on A Molecular Phylogeny of the Pheasants and Partridges Suggests That These Lineages Are Not Monophyletic R. T. Kimball,* E. L. Braun,*,† P. W. Zwartjes,* T. M. Crowe,‡,§ and J. D. Ligon* *Department of Biology, University of New Mexico, Albuquerque, New Mexico 87131; †National Center for Genome Resources, 1800 Old Pecos Trail, Santa Fe, New Mexico 87505; ‡Percy FitzPatrick Institute, University of Capetown, Rondebosch, 7700, South Africa; and §Department of Ornithology, American Museum of Natural History, Central Park West at 79th Street, New York, New York 10024-5192 Received October 8, 1997; revised June 2, 1998 World partridges are smaller and widely distributed in Cytochrome b and D-loop nucleotide sequences were Asia, Africa, and Europe. Most partridge species are used to study patterns of molecular evolution and monochromatic and primarily dull colored. None exhib- phylogenetic relationships between the pheasants and its the extreme or highly specialized ornamentation the partridges, which are thought to form two closely characteristic of the pheasants. related monophyletic galliform lineages. Our analyses Although the order Galliformes is well defined, taxo- used 34 complete cytochrome b and 22 partial D-loop nomic relationships are less clear within the group sequences from the hypervariable domain I of the (Verheyen, 1956), due to the low variability in anatomi- D-loop, representing 20 pheasant species (15 genera) and 12 partridge species (5 genera). We performed cal and osteological traits (Blanchard, 1857, cited in parsimony, maximum likelihood, and distance analy- Verheyen, 1956; Lowe, 1938; Delacour, 1977). -

A Baraminological Analysis of the Land Fowl (Class Aves, Order Galliformes)
Galliform Baraminology 1 Running Head: GALLIFORM BARAMINOLOGY A Baraminological Analysis of the Land Fowl (Class Aves, Order Galliformes) Michelle McConnachie A Senior Thesis submitted in partial fulfillment of the requirements for graduation in the Honors Program Liberty University Spring 2007 Galliform Baraminology 2 Acceptance of Senior Honors Thesis This Senior Honors Thesis is accepted in partial fulfillment of the requirements for graduation from the Honors Program of Liberty University. ______________________________ Timothy R. Brophy, Ph.D. Chairman of Thesis ______________________________ Marcus R. Ross, Ph.D. Committee Member ______________________________ Harvey D. Hartman, Th.D. Committee Member ______________________________ Judy R. Sandlin, Ph.D. Assistant Honors Program Director ______________________________ Date Galliform Baraminology 3 Acknowledgements I would like to thank my Lord and Savior, Jesus Christ, without Whom I would not have had the opportunity of being at this institution or producing this thesis. I would also like to thank my entire committee including Dr. Timothy Brophy, Dr. Marcus Ross, Dr. Harvey Hartman, and Dr. Judy Sandlin. I would especially like to thank Dr. Brophy who patiently guided me through the entire research and writing process and put in many hours working with me on this thesis. Finally, I would like to thank my family for their interest in this project and Robby Mullis for his constant encouragement. Galliform Baraminology 4 Abstract This study investigates the number of galliform bird holobaramins. Criteria used to determine the members of any given holobaramin included a biblical word analysis, statistical baraminology, and hybridization. The biblical search yielded limited biosystematic information; however, since it is a necessary and useful part of baraminology research it is both included and discussed. -

Hybridization & Zoogeographic Patterns in Pheasants
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Paul Johnsgard Collection Papers in the Biological Sciences 1983 Hybridization & Zoogeographic Patterns in Pheasants Paul A. Johnsgard University of Nebraska-Lincoln, [email protected] Follow this and additional works at: https://digitalcommons.unl.edu/johnsgard Part of the Ornithology Commons Johnsgard, Paul A., "Hybridization & Zoogeographic Patterns in Pheasants" (1983). Paul Johnsgard Collection. 17. https://digitalcommons.unl.edu/johnsgard/17 This Article is brought to you for free and open access by the Papers in the Biological Sciences at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Paul Johnsgard Collection by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. HYBRIDIZATION & ZOOGEOGRAPHIC PATTERNS IN PHEASANTS PAUL A. JOHNSGARD The purpose of this paper is to infonn members of the W.P.A. of an unusual scientific use of the extent and significance of hybridization among pheasants (tribe Phasianini in the proposed classification of Johnsgard~ 1973). This has occasionally occurred naturally, as for example between such locally sympatric species pairs as the kalij (Lophura leucol11elana) and the silver pheasant (L. nycthelnera), but usually occurs "'accidentally" in captive birds, especially in the absence of conspecific mates. Rarely has it been specifically planned for scientific purposes, such as for obtaining genetic, morphological, or biochemical information on hybrid haemoglobins (Brush. 1967), trans ferins (Crozier, 1967), or immunoelectrophoretic comparisons of blood sera (Sato, Ishi and HiraI, 1967). The literature has been summarized by Gray (1958), Delacour (1977), and Rutgers and Norris (1970). Some of these alleged hybrids, especially those not involving other Galliformes, were inadequately doculnented, and in a few cases such as a supposed hybrid between domestic fowl (Gallus gal/us) and the lyrebird (Menura novaehollandiae) can be discounted. -

Assessment of Hematological Indices of Indian Peafowl (Pavo Cristatus) Kept at Wildlife Breeding Centre, Gatwala, Faisalabad, Pakistan
Journal of Zoological Research Volume 4, Issue 1, 2020, PP 29-33 ISSN 2637-5575 Assessment of Hematological Indices of Indian Peafowl (Pavo Cristatus) Kept at Wildlife Breeding Centre, Gatwala, Faisalabad, Pakistan Misbah Sarwar1* Zahid Ali1, and Muhammad Bilal Chaudhary2 1Punjab Wildlife Research Centre, Gatwala, Faisalabad Department of Wildlife & Parks, Punjab Pakistan 2Department of Zoology, GC University, Faisalabad, Pakistan *Corresponding Author: Misbah Sarwar, Punjab Wildlife Research Centre, Gatwala, Faisalabad Department of Wildlife & Parks, Punjab Pakistan. Email: [email protected] ABSTRACT Indian peafowl (Pavo cristatus) or blue peafowl has been maintained in captivity since long where due to selective breeding, several color mutations/varieties have appeared of which white peafowl, black-shouldered peafowl and pied peafowl are common. Since, hematological analysis is crucial for clinical diagnosis of wild and captive avifauna, so we collected blood samples from healthy male blue peafowl, white peafowl and black- shouldered peafowl kept at Wildlife Breeding Centre, Gatwala, Faisalabad and compared erythrocyte and leucocyte indices among them. Our results indicated that blood physiological values for MO (%), Hgb, HCT, MCH and MCHC were significantly different (P<0.05) between blue peafowl and white peafowl whereas MCV and RDW were significantly different (P<0.05) between blue peafowl and black-shouldered peafowl. The comparison of hematological parameters between white peafowl and black-shouldered peafowl showed that GR(%), RBC, HCT, MCV and MCHC differ significantly (P<0.05) between the two varieties. Our results support the studies indicating high quality color patterns reflect increased resistance and immunity to pathogens. Keywords: Indian peafowl, Color Mutations, hematology. INTRODUCTION 2005; Takahashi and Hasegawa, 2008; Harikrishnan et al., 2010, Naseer et al., 2017). -

Peafowl in Spring, Peacocks Begin Establishing Leks, Incubation Period Begins
may be impressed with the peacock’s beauty, Peahens conceal their nests, a simple scrape Visitor Centers & Recreation Services the true target of all his glamorous excess is lined with grass, in thick vegetation. The eggs ARDENWOOD HISTORIC FARM his potential mate, the peahen. are laid every other day until a total of up to Fremont 510-796-0199, [email protected] eight is reached, at which point the 28 day Peafowl In spring, peacocks begin establishing leks, incubation period begins. The peachicks are BLACK DIAMOND MINES a group of small adjacent territories, each of “precocial,” meaning they are born with their Antioch 925-757-2620, [email protected] which is the domain of a single male. Inter- eyes open, feathered, and ready to walk soon ested females visit the lek, assessing the vari- COYOTE HILLS REGIONAL PARK after hatching. Peahens are attentive mothers, Fremont 510-795-9385, [email protected] ous males before choosing a mate. In order but the peacock is not involved in raising his to attract mates, the peacock employs his young. The next generation of peahens will be CRAB COVE at CROWN BEACH most striking feature, the breathtaking train ready to reproduce at one or two years of age Alameda 510-521-6887, [email protected] of feathers often referred to as the peacock’s while the males need three years to develop tail. This 4 foot long collection of hundreds SUNOL REGIONAL WILDERNESS a full train of display plumage. Peafowl are Sunol 925-862-2601, [email protected] of feathers is actually made up of the upper hardy birds and may live 20 years or more. -
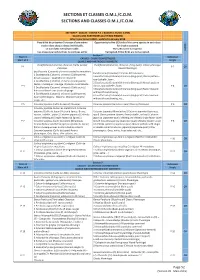
Section P, Quales and Partridges.Cdr
SECTIONS ET CLASSES O.M.J./C.O.M. SECTIONS AND CLASSES O.M.J./C.O.M. SECTION P - CAILLES - COLINS P.E. ( BAGUES 1 ET/OU 2 ANS) QUAILS AND PARTRIDGES (1/2 YEARS RINGED) Mise à jour Janvier 2018 - updated in January 2018 Possibilité de présenter 5 oiseaux d'une même Opportunity to five (5) birds of the same species in each class C A espèce dans chaque classe individuelle. for singles accepted GE Le nom lan est indispensable. The Lan name is required. S Les oiseaux panachés-frisés ne sont pas admis Variegated-frilled birds are not accepted. Secon P Stam 4 Individuel CAILLES - COLINS DOMESTIQUES Stam of 4 Single QUAILS AND PARTRIDGES DOMESTIC Excalfactoria (Coturnix) chinensis (Caille peinte) Excalfactoria(coturnix) chinensis. (King quail) Classic phenoype P 1 P 2 O1 Classique Classic Phenotype Excalfactoria (Coturnix) chinensis toutes les mutaons Excalfactoria (Coturnix) chinensis All mutaons: 1 Excalfactoria (Coturnix) chinensis (Caille peinte) 1 Excalfactoria (Coturnix) chinensis (King quail) Classic paern : Dessin sauvage : Opale,Brune etIsabelle . opal,Isabelle , fawn.. 2 Excalfactoria (Coturnix) chinensis (Caille peinte) 2 Excalfactoria (Coturnix) chinensis (King quail) Mosaïc paern Facteur mosaïque : sauvage, Opale,Brune etIsabelle P 3 classic, opal,Isabelle , fawn. P 4 O1 3 Excalfactoria (Coturnix) chinensis (Caille peinte) 3 Excalfactoria (Coturnix) chinensis (King quail) Factor melanic Patron mélanisé sans dessin de gorge without throat drawing 4 Excalfactoria (Coturnix) chinensis (Caille peinte) 4 Excalfactoria (Coturnix) chinensis -

Comparison of Cytochrome B Region Among Chinese Painted Quail, Wild-Strain Quail, and White Broiler Chicken Based on PCR-RFLP Analysis
287 Comparison of Cytochrome b Region Among Chinese Painted Quail, Wild-Strain Quail, and White Broiler Chicken Based on PCR-RFLP Analysis Xiang-Jun SHEN1),Hidenori SUZUKI2)*, Masaoki TSUDZUKI3), Shin'ichiITO2) and Takao NAKAMURA2)** 1)The United Graduate School of AgriculturalScience , 2)Faculty of Agriculture, GifuUniversity, Gifu 501-1193 3)Faculty of AppliedBiological Science , Hiroshima University,Higashi-Hiroshima 739-8528 PCR-RFLPanalysis was used to examine 1,075 bp cytochromeb (Cyt b) region of mtDNAineach of 20birds of Chinese painted quail (Excalfactoria chinensis), wild-strain Japanesequail (Coturnix japonica), and white broiler chicken (Gallus gallus). The total Cytb ampliconwas digested with 10 restriction endonucleases, andits electrophoretic patternwas investigated. Ten kinds of restrictionenzyme digestions were identical among20 individuals ineach of the three species. No variation was observed in the 10 kindsof enzymedigestions among different individuals within the threespecies in- vestigated.The different specific electrophoretic patterns, haplotypes, and total restric- tionfragments were observed in eachof the three species belonging tothe same family Phasianidae.The result representatives the evolutionarygenetic characteristics of RFLPof cytochromeb region in thethree different species, and indicated the genetic diversityamong the three genuses of the family Phasianidae, and genetic identity within eachof thespecies. The result could be used for the identification of different species withinfamily Phasianidae and as a referenceofphysical map for cytochrome b gene. (Jpn.Poult. Sci., 36: 287-294, 1999) Keywords:PCR-RFLP, cytochrome b,Excalfactoria chinensis, Coturnix japonica, Gallus gallus Introduction Chinesepainted quail (Excalfactoria chinensis) belongs to the orderGalliformes, family Phasianidae,and genus Excalfactoria,(YAMASHINA, 1986). Japanese quail belongsto the samefamily Phasianidae, a different genus Coturnix (CRAWFORD, 1990). -

Distribution and Conservation Status of Congo Peafowl Afropavo Congensis in Eastern Zaire
Bird Conservation International (1997) 7:295-316. © BirdLife International 1997 Distribution and conservation status of Congo Peafowl Afropavo congensis in eastern Zaire JOHN A. HART and AGENOGA UPOKI Summary Between 1993 and 1995 the occurrence and status of Congo Peafowl Afropavo congensis were assessed at 89 sites in eastern Zaire based on interviews with local hunters (114 locations) and forest surveys (24 locations). The survey region covered approximately 125,000 km2 and was bounded by the Lomami River (24° 30') to the west, the Albertine Rift Highlands (290) to the east, the Aruwimi River (20 N) to the north and the Kahuzi Biega Park (30 S) to the south. Congo Peafowl have been extirpated or were only doubtfully present at 16 of 65 sites where survey coverage was adequate and where the species was reported to have occurred in the past. Occurrence of the species was confirmed or probable at the remaining 49 sites. Congo Peafowl were reported as locally common (> five reported encounters since 1990) at only 12 sites. We observed or heard peafowl at five sites. A single nest was found which contained two eggs. Observations on group size and habitat associations were also made. Congo Peafowl were considered to be seriously threatened at 19 of the 65 sites. Captures in snares set for small mammals and antelope are a major threat to Congo Peafowl throughout most of the survey region. Mining, shifting cultivation and logging leading to habitat loss were significant threats in several locations. Congo Peafowl appear to have disappeared from several sites where human activity was frequent, but habitat loss was not extensive. -

Rapid Communication- Primer Design and Sequence Information for Cytochrome B Gene in the Chinese Painted Quail (Excalfactoria C
-Rapid Communication- Primer Design and Sequence Information for Cytochrome b Gene in the Chinese Painted Quail (Excalfactoria chinensis) Xiang-Jun SHEN, Masaoki TSUDZUKIa, Takao NAKAMURAb The United Graduate School of Agricultural Science, Gifu University, Gifu-shi 501-1193, Japan Faculty of Applied Biological Science, Hiroshima University, Higashi-Hiroshima-shi 739-8528,a Japan bFaculty of Agriculture , Gifu University, Gifu-shi 501-1193, Japan (Received April 21, 1999; Accepted May 15, 1999) Abstract In accordance with the chicken and quail mitochondrialDNA sequences,three pairs of primers encompassing light and heavy strand (L14832, L15269, L15500, H15924, H15497, and H16207) with high specificity were designed, synthesized, and used for amplification and sequencing of cytochrome b (Cyt b) gene. The PCR products were sequenced by the direct-sequencing method. The Chinese painted quail cytochrome b gene consisted of 1,143bp nucleotides (base composition: 327 A; 399 C; 281 T and 136 G), encoded for a 380 amino acid sequence, and showed 89.0%, 86.4%, and 86.2%, homology with nucleotide sequences of Japanese quail, silky fowl, and White Leghorn chicken, respectively. The homology of the amino acid sequence of the cytochrome b gene of Chinese painted quail showed 98.2%, 95.3%, and 95.3% with those of Japanese quail, silky fowl, and White Leghorn chicken, respectively. The sequencesreported in this paper have been depositedin the GenBank database(Accession No. AF109217). Animal Science Journal 70 (4): 240-242,1999 Key words: Chinese painted quail,Excalfactoria chinensis, Cytochromeb, Sequence,Homology Chinese painted quail (Excalfactoria chinensis) belongs the test: to the order Galliformes, family Phasianidae, genus (1) L14832: 5'-TACCTGGGTTCCTTCGCCCT-3' Excalfactoria5),and it is the smallest species in the family (2) H15497: 5'-TGTAGGAAGGTGAGTGGAT-3' Phasianidae. -

The Status and Distribution of Green Peafowl Pavo Muticus in Yunnan Province, China
The status and distribution of green peafowl Pavo muticus in Yunnan Province, China LIANXIAN HAN *, YUEQIANG LIU and BENG HAN Faculty of Conservation Biology, Southwest Forestry College, Kunming Yunnan China 650224. *Correspondence author - [email protected] Paper presented at the 4 th International Galliformes Symposium, 2007, Chengdu, China. Abstract Data on the status and distribution of the green peafowl in Yunnan Province, China, were collected from historical data, field surveys carried out between March and June 2007 around Shuangbai and Baoshan, and from a correspondence survey among city, state and county Forestry Bureaux and Protection Agencies. The data showed that green peafowl occurred historically in at least 42 locations, but by the late 1980s they were already absent from Yingjiang, Tengchong, Liuku, Mengzi and Hekou, and by the late 1990s were also absent from Mengla, Jinping and Luchun and Jianshui County. Green peafowl were found to be present in 31 known areas: Weishan, Yongren, Jinghong, Ruili, Longchuan, Luxi, Longling, Changning, Fengqing, Yunxian, Yongde, Chenkang, Kengma, Cangyuan, Shuangjiang, Lincang, Jingdong, Jinggu, Chenyuan, Pu'er, Simao, Menghai, Mojiang, Shiping, Maitreya, Sinping, Shuangbai, Chuxiong, Lufeng, Nanhua, and Yao'an. Among these, Weishan, Yongren, Jing Hong, Mengla have only supported green peafowl since the latter half of the 1980s. This evaluation also located green peafowl in six new areas, namely Baoshan, Nanjian, Lancang, Yuxi, Shidian and Honghe. It remains unknown if two further -

Testing Darwin's Hypothesis on the Evolution of Ornamental Eyespots In
Testing Darwin’s hypothesis on the evolution of ornamental eyespots in peafowl and their relatives Rebecca T. Kimball, Keping Sun, Kelly Meicklejohn, Edward Braun, Brant Faircloth, Travis Glenn Darwin • “As no ornaments are more beautiful than the ocelli on the feather of various birds…they deserve to be especially noticed.” (The Descent of Man: And Selection in Relation to Sex) Hypothesis • Darwin suggested a gradual evolution from the simple spots and ocelli of some species, to the magnificent ocelli of peafowl Ocelli in other taxa • Noted similarities between peafowl (Pavo), argus pheasant (Argusianus), and peacock pheasants (Polyplectron) Relationships • Based on the shared ocelli, some classifications have united the peafowl, peacock pheasants and argus pheasants into a clade • However, molecular phylogenies have not been able to resolve relationships of these taxa with confidence What we do know • Based on molecular phylogenies: – The peafowl are most closely related to the Congo peafowl (which lacks ocelli) – The argus pheasant is most closely related to the crested argus (which lacks ocelli) • So the simplest interpretation of Darwin’s ideas, that the ocellated taxa would be each other’s closest relatives, is not supported Ocelli can be lost • Within the peacock pheasants, one species lacks complex ocelli – This appears to be a derived species – So loss of ocelli does occur • A modification of Darwin’s hypothesis might be that ocelli evolved once, but have been lost in Congo peacock, crested argus, and one species of peacock pheasant