Statistical Data Integration in Translational Medicine
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Functional Effects Detailed Research Plan
GeCIP Detailed Research Plan Form Background The Genomics England Clinical Interpretation Partnership (GeCIP) brings together researchers, clinicians and trainees from both academia and the NHS to analyse, refine and make new discoveries from the data from the 100,000 Genomes Project. The aims of the partnerships are: 1. To optimise: • clinical data and sample collection • clinical reporting • data validation and interpretation. 2. To improve understanding of the implications of genomic findings and improve the accuracy and reliability of information fed back to patients. To add to knowledge of the genetic basis of disease. 3. To provide a sustainable thriving training environment. The initial wave of GeCIP domains was announced in June 2015 following a first round of applications in January 2015. On the 18th June 2015 we invited the inaugurated GeCIP domains to develop more detailed research plans working closely with Genomics England. These will be used to ensure that the plans are complimentary and add real value across the GeCIP portfolio and address the aims and objectives of the 100,000 Genomes Project. They will be shared with the MRC, Wellcome Trust, NIHR and Cancer Research UK as existing members of the GeCIP Board to give advance warning and manage funding requests to maximise the funds available to each domain. However, formal applications will then be required to be submitted to individual funders. They will allow Genomics England to plan shared core analyses and the required research and computing infrastructure to support the proposed research. They will also form the basis of assessment by the Project’s Access Review Committee, to permit access to data. -

Algorithms for Computational Biology 8Th International Conference, Alcob 2021 Missoula, MT, USA, June 7–11, 2021 Proceedings
Lecture Notes in Bioinformatics 12715 Subseries of Lecture Notes in Computer Science Series Editors Sorin Istrail Brown University, Providence, RI, USA Pavel Pevzner University of California, San Diego, CA, USA Michael Waterman University of Southern California, Los Angeles, CA, USA Editorial Board Members Søren Brunak Technical University of Denmark, Kongens Lyngby, Denmark Mikhail S. Gelfand IITP, Research and Training Center on Bioinformatics, Moscow, Russia Thomas Lengauer Max Planck Institute for Informatics, Saarbrücken, Germany Satoru Miyano University of Tokyo, Tokyo, Japan Eugene Myers Max Planck Institute of Molecular Cell Biology and Genetics, Dresden, Germany Marie-France Sagot Université Lyon 1, Villeurbanne, France David Sankoff University of Ottawa, Ottawa, Canada Ron Shamir Tel Aviv University, Ramat Aviv, Tel Aviv, Israel Terry Speed Walter and Eliza Hall Institute of Medical Research, Melbourne, VIC, Australia Martin Vingron Max Planck Institute for Molecular Genetics, Berlin, Germany W. Eric Wong University of Texas at Dallas, Richardson, TX, USA More information about this subseries at http://www.springer.com/series/5381 Carlos Martín-Vide • Miguel A. Vega-Rodríguez • Travis Wheeler (Eds.) Algorithms for Computational Biology 8th International Conference, AlCoB 2021 Missoula, MT, USA, June 7–11, 2021 Proceedings 123 Editors Carlos Martín-Vide Miguel A. Vega-Rodríguez Rovira i Virgili University University of Extremadura Tarragona, Spain Cáceres, Spain Travis Wheeler University of Montana Missoula, MT, USA ISSN 0302-9743 ISSN 1611-3349 (electronic) Lecture Notes in Bioinformatics ISBN 978-3-030-74431-1 ISBN 978-3-030-74432-8 (eBook) https://doi.org/10.1007/978-3-030-74432-8 LNCS Sublibrary: SL8 – Bioinformatics © Springer Nature Switzerland AG 2021 This work is subject to copyright. -

Female Fellows of the Royal Society
Female Fellows of the Royal Society Professor Jan Anderson FRS [1996] Professor Ruth Lynden-Bell FRS [2006] Professor Judith Armitage FRS [2013] Dr Mary Lyon FRS [1973] Professor Frances Ashcroft FMedSci FRS [1999] Professor Georgina Mace CBE FRS [2002] Professor Gillian Bates FMedSci FRS [2007] Professor Trudy Mackay FRS [2006] Professor Jean Beggs CBE FRS [1998] Professor Enid MacRobbie FRS [1991] Dame Jocelyn Bell Burnell DBE FRS [2003] Dr Philippa Marrack FMedSci FRS [1997] Dame Valerie Beral DBE FMedSci FRS [2006] Professor Dusa McDuff FRS [1994] Dr Mariann Bienz FMedSci FRS [2003] Professor Angela McLean FRS [2009] Professor Elizabeth Blackburn AC FRS [1992] Professor Anne Mills FMedSci FRS [2013] Professor Andrea Brand FMedSci FRS [2010] Professor Brenda Milner CC FRS [1979] Professor Eleanor Burbidge FRS [1964] Dr Anne O'Garra FMedSci FRS [2008] Professor Eleanor Campbell FRS [2010] Dame Bridget Ogilvie AC DBE FMedSci FRS [2003] Professor Doreen Cantrell FMedSci FRS [2011] Baroness Onora O'Neill * CBE FBA FMedSci FRS [2007] Professor Lorna Casselton CBE FRS [1999] Dame Linda Partridge DBE FMedSci FRS [1996] Professor Deborah Charlesworth FRS [2005] Dr Barbara Pearse FRS [1988] Professor Jennifer Clack FRS [2009] Professor Fiona Powrie FRS [2011] Professor Nicola Clayton FRS [2010] Professor Susan Rees FRS [2002] Professor Suzanne Cory AC FRS [1992] Professor Daniela Rhodes FRS [2007] Dame Kay Davies DBE FMedSci FRS [2003] Professor Elizabeth Robertson FRS [2003] Professor Caroline Dean OBE FRS [2004] Dame Carol Robinson DBE FMedSci -
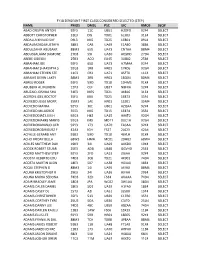
FY18 SFC AC Considered Selected.Xlsx
FY18 SERGEANT FIRST CLASS CONSIDERED SELECTED (STEP) NAME PMOS DMSL PSC UIC RMOS SEQ# ABAD CRISPIN ANTON 92F3 11C UB51 H2DFD 92F4 SELECT ABBOTT CHRISTOPHER 31E3 DIS TD01 6L303 31E4 SELECT ABDALLA KHALID SAF 09L3 MIS TD25 1E801 09L4 SELECT ABDALRAZAQ MUSTAFA 38B3 CAB UA69 E1AB0 38B4 SELECT ABDULZAHIR ABUBAKR 88M3 6SG UA73 CNTAA 88M4 SELECT ABDUSSALAAM DAMONE 27D3 S3I UA30 AQSHD 27D4 SELECT ABEBE GIDEON 25B3 ACO EU45 3U802 25B4 SELECT ABRAHAM JOE 92F3 6SG UA73 H7MAA 92F4 SELECT ABRAHAM SHAKEMYA S 92G3 1RB HR01 17R06 92G4 SELECT ABRAHAM STEVEN STE 11C3 CR2 UA71 ASFT0 11C4 SELECT ABRAMS DEVIN LAKEY 88M3 2RB HR01 18Q04 88M4 SELECT ABREU ROGER 91F3 59O TD18 1D4M1 91X4 SELECT ABUBEKR ALI NURDIN 12P3 CEF UB37 NBFXA 12P4 SELECT ABUZAID JOSHUA SHA 31E3 MPS TD21 4K816 31E4 SELECT ACERON JOEL BOCTOT 35F3 MIS TD25 1E873 35F4 SELECT ACEVEDO JESSE MORR 35M3 1AE HR01 1S201 35M4 SELECT ACEVEDO MAYRA 92Y3 36E UB01 AZQAA 92Y4 SELECT ACEVEDO MILAGROS 35S3 MIS TD15 1E874 35S4 SELECT ACEVEDODIAZ LUIS H 92G3 H82 UA15 AA6T0 92G4 SELECT ACEVEDOMARRS MARYS 92G3 IMS MD11 0UC1A 92G4 SELECT ACEVEDONARANJO GER 92Y3 173 UA70 ELGAA 92Y4 SELECT ACEVEDORODRIGUEZ F 42A3 IGH FS27 2L623 42A4 SELECT ACFALLE GERARD ANT 91B3 59O TD18 46YLH 91X4 SELECT ACHO FRIDAY BELLA 68W3 HMH MC01 2DH03 68W4 SELECT ACKLES MATTHEW DAV 19K3 34I UA93 AX6D0 19K4 SELECT ACOCK ROBERT STUAR 25E3 ADB UB48 GGVHD 25E4 SELECT ACORD MATTHEW STEP 92Y3 2FO UA53 JX6AA 92Y4 SELECT ACOSTA ALBERTO EDU 74D3 3CB TD21 4K901 74D4 SELECT ACOSTA MARTIN ALON 18E3 S07 UA38 H01A0 18E4 SELECT ACQUI STEPHEN D 88M3 14I -

Beendigung Der Entzündungsreaktion Durch Interleukin-9 Sezernierende
Beendigung der Entz¨undungsreaktion durch Interleukin-9 sezernierende angeborene Lymphozyten Der Naturwissenschaftlichen Fakult¨at der Friedrich-Alexander-Universit¨at Erlangen-Nurnberg¨ zur Erlangung des Doktorgrades Dr. rer. nat. vorgelegt von Simon Rauber Als Dissertation genehmigt von der Naturwissenschaftlichen Fakult¨at der Friedrich-Alexander-Universit¨at Erlangen-Nurnberg¨ Tag der mundlichen¨ Prufung:¨ 02. 05. 2919 Vorsitzender des Promotionsorgans: Prof. Dr. Georg Kreimer Gutachter: Prof. Dr. Steffen Backert Prof. Dr. Georg Schett Resolution of inflammation by interleukin-9 producing innate lymphoid cells To the Faculty of Natural Sciences of the Friedrich-Alexander-University Erlangen-Nuremberg for the obtainment of the academic degree doctor rerum naturalium (Dr. rer. nat.) submitted by Simon Rauber Approved by the Faculty of Natural Sciences of the Friedrich-Alexander-University Erlangen-Nuremberg Date of oral examination: 02. 05. 2019 Chairman of examination board: Prof. Dr. Georg Kreimer Referees: Prof. Dr. Steffen Backert Prof. Dr. Georg Schett Table of contents 1 Deutsche Kurzfassung1 2 English abstract3 3 Introduction 5 3.1 Innate lymphoid cells and the bridge between primary and adaptive immune response6 3.1.1 The discovery . .7 3.1.2 The classification of innate lymphoid cells and their integration into the im- mune system . 11 3.1.3 Are innate lymphoid cells only innate mirrors or fully-fledged immune cells . 12 3.2 The interleukin-9 . 15 3.2.1 The cellular source of interleukin-9 . 15 3.2.2 The immune-modulatory capacities of interleukin-9 . 17 3.2.2.1 Interleukin-9 in infection, tumour and allergy . 18 3.2.2.2 Interleukin-9 in autoimmune diseases . -

Autoantibody-Mediated Arthritis in The
Auger et al. Arthritis Research & Therapy 2012, 14:R269 http://arthritis-research.com/content/14/6/R269 RESEARCHARTICLE Open Access Autoantibody-mediated arthritis in the absence of C3 and activating Fcg receptors: C5 is activated by the coagulation cascade Jennifer L Auger1, Stefanie Haasken1,2 and Bryce A Binstadt1* Abstract Introduction: The effector functions of immunoglobulin G (IgG) are mediated by interaction of its Fc region with Fc receptors (FcgRs) and/or the complement system. The three main pathways of complement activation converge at C3. However, C3-independent pathways can activate C5 and other downstream complement components during IgG-initiated inflammatory responses. These C3-independent pathways of C5 activation are triggered by activating FcgRs in some systems or can be activated by factors of the coagulation cascade such as thrombin. Here we studied the interplay of C3, C5, and activating FcgRs in a model of spontaneous autoantibody-driven arthritis. Methods: We utilized the K/BxN TCR transgenic mouse model of arthritis. We bred K/BxN mice bearing targeted or naturally-occurring mutations in one or more of the genes encoding complement components C3, C5, and FcRg, the cytoplasmic signaling chain shared by the activating FcgRs. We measured arthritis development, the production of arthritogenic autoantibodies, T cell activation status and cytokine synthesis. In addition, we treated mice with anti-C5 monoclonal antibodies or with the thrombin inhibitor argatroban. Results: We have previously shown that genetic deficiency of C5 protects K/BxN mice from the development of arthritis. We found here that C3-deficient K/BxN mice developed arthritis equivalent in severity to C3-sufficient animals. -

Computational Biology and Bioinformatics
Vol. 30 ISMB 2014, pages i1–i2 BIOINFORMATICS EDITORIAL doi:10.1093/bioinformatics/btu304 Editorial This special issue of Bioinformatics serves as the proceedings of The conference used a two-tier review system, a continuation the 22nd annual meeting of Intelligent Systems for Molecular and refinement of a process begun with ISMB 2013 in an effort Biology (ISMB), which took place in Boston, MA, July 11–15, to better ensure thorough and fair reviewing. Under the revised 2014 (http://www.iscb.org/ismbeccb2014). The official confer- process, each of the 191 submissions was first reviewed by at least ence of the International Society for Computational Biology three expert referees, with a subset receiving between four and (http://www.iscb.org/), ISMB, was accompanied by 12 Special eight reviews, as needed. These formal reviews were frequently Interest Group meetings of one or two days each, two satellite supplemented by online discussion among reviewers and Area meetings, a High School Teachers Workshop and two half-day Chairs to resolve points of dispute and reach a consensus on tutorials. Since its inception, ISMB has grown to be the largest each paper. Among the 191 submissions, 29 were conditionally international conference in computational biology and bioinfor- accepted for publication directly from the first round review Downloaded from matics. It is expected to be the premiere forum in the field for based on an assessment of the reviewers that the paper was presenting new research results, disseminating methods and tech- clearly above par for the conference. A subset of 16 papers niques and facilitating discussions among leading researchers, were viewed as potentially in the top tier but raised significant practitioners and students in the field. -

Veterans Memorial (By Names & Numbers).Xlsx
Name Service Branch Conflict Monument ID# Entries Emblem at Top of Mon. Abbott, Donald H. US Army WWII 18-B-Right Col 9 3632 Disable American Veterans Abbott, James P. US Air Force Persian Gulf 18-B-Right Col 23 3646 Disable American Veterans Abbott, John D. US Air Force Vietnam 18-B-Right Col 21 3644 Disable American Veterans Abbott, John D. Jr. US Air Force Panama 18-B-Right Col 22 3645 Disable American Veterans Abbott, Robert P. US Marine Corps Vietnam 28-B-Left Col. 19 5748 Air National Guard Achorn, Richard L. US Navy WWII 8-A Right Col. 46 1612 POW-MIA Logo Adamen, Stanley M. US Army WWII 17-B-Left Col 14 3367 AMVETS Adams, Bernard L. Jr. US Army WWII 28-A-Left Col. 14 5635 Air National Guard Adams, Byron C. Jr. US Army WWII 11-B-Right Col. 44 2366 Veterans of Foreign Wars Adams, Garfield D. US Navy WWII 20-A-Right Col 10 3957 US Army WAC Logo Adams, Harold C. US Army WWII 22-A-Left Col 15 4340 US Navy Waves Adams, Harold F. US Army Vietnam 32-A-Right Col. 21 6560 Destroyer Escort Assoc. Adams, Preston G. US Navy WWII 21-A-Left Col 24 4133 US Air Force Women Adams, Ralph S. US Army Korea 2-B Right Col. 33 411 State of Maine Seal Adams, Richard C. US Army WWI 22-B-Left Col 11 4444 US Navy Waves Adams, Richard M. US Army WWII 10-B Right Col. 40 2146 Order of the Purple Heart Adams, Robert P. -

Cancer Immunology of Cutaneous Melanoma: a Systems Biology Approach
Cancer Immunology of Cutaneous Melanoma: A Systems Biology Approach Mindy Stephania De Los Ángeles Muñoz Miranda Doctoral Thesis presented to Bioinformatics Graduate Program at Institute of Mathematics and Statistics of University of São Paulo to obtain Doctor of Science degree Concentration Area: Bioinformatics Supervisor: Prof. Dr. Helder Takashi Imoto Nakaya During the project development the author received funding from CAPES São Paulo, September 2020 Mindy Stephania De Los Ángeles Muñoz Miranda Imunologia do Câncer de Melanoma Cutâneo: Uma abordagem de Biologia de Sistemas VERSÃO CORRIGIDA Esta versão de tese contém as correções e alterações sugeridas pela Comissão Julgadora durante a defesa da versão original do trabalho, realizada em 28/09/2020. Uma cópia da versão original está disponível no Instituto de Matemática e Estatística da Universidade de São Paulo. This thesis version contains the corrections and changes suggested by the Committee during the defense of the original version of the work presented on 09/28/2020. A copy of the original version is available at Institute of Mathematics and Statistics at the University of São Paulo. Comissão Julgadora: • Prof. Dr. Helder Takashi Imoto Nakaya (Orientador, Não Votante) - FCF-USP • Prof. Dr. André Fujita - IME-USP • Profa. Dra. Patricia Abrão Possik - INCA-Externo • Profa. Dra. Ana Carolina Tahira - I.Butantan-Externo i FICHA CATALOGRÁFICA Muñoz Miranda, Mindy StephaniaFicha de Catalográfica los Ángeles M967 Cancer immunology of cutaneous melanoma: a systems biology approach = Imuno- logia do câncer de melanoma cutâneo: uma abordagem de biologia de sistemas / Mindy Stephania de los Ángeles Muñoz Miranda, [orientador] Helder Takashi Imoto Nakaya. São Paulo : 2020. 58 p. -

Clov) (Seafarer
REPUBLIC OF THE PHILIPPINES COMMISSIONS ON ELECTIONS OFFICE FOR OVERSEAS VOTING CERTIFIED LIST OF OVERSEAS VOTERS (CLOV) (SEAFARER) Source: Server Seq No. Voter's Name REGISTRATION DATE 1 ABA, REGINALD PATRON September 28, 2014 2 ABAA, ADRIAN SIMBORIO February 26, 2015 3 ABABA, JOVITO SUMAYAO October 03, 2012 4 ABABON, MELECIO VARGA October 10, 2014 5 ABACIAL, JEAM URIEL BOHOS March 26, 2018 6 ABAD, ALDIN BIANZON August 07, 2014 7 ABAD, ALDRICH BRAVO November 10, 2014 8 ABAD, ALFONSO JR. LOPEZ September 18, 2015 9 ABAD, ANABELLE NAMBAYAN September 16, 2015 10 ABAD, ANTONIO JR. VASQUES August 28, 2015 11 ABAD, AUGUSTO DIZON February 11, 2015 12 ABAD, BRIAN LAWAY October 26, 2015 13 ABAD, CARLA MICHELLE PUNIO May 07, 2015 14 ABAD, CESAR JR. ORALLO July 07, 2015 15 ABAD, DIGO BETUA January 28, 2015 16 ABAD, EDGARDO BUSA September 13, 2012 17 ABAD, EDUWARDO DE LUMEN September 24, 2018 18 ABAD, ERNESTO ESMAO August 11, 2015 19 ABAD, ERWIN TRINOS May 06, 2006 20 ABAD, FERDINAND BENTULAN August 07, 2014 21 ABAD, GERALD QUIL MAPANAO August 25, 2015 22 ABAD, HECTOR PASION August 05, 2015 23 ABAD, HERMIE MANALO August 28, 2015 24 ABAD, JEFFREY BRAVO May 25, 2018 25 ABAD, JOEL FRANCIA September 29, 2015 26 ABAD, JOEL RODRIGUEZ October 05, 2006 27 ABAD, JOHN CHESTER ABELLA August 05, 2015 28 ABAD, MARLON EVANGELISTA November 16, 2017 29 ABAD, NERIE GERSALIA January 17, 2018 30 ABAD, NOLI SOMIDO October 13, 2015 31 ABAD, PATRICK GACULA January 24, 2018 32 ABAD, RICKY MOJECA September 21, 2018 33 ABAD, ROMULO CERVANTES April 20, 2012 34 ABAD, RONALD TAN May 08, 2017 35 ABADAM, ALAN BASILIA August 19, 2012 NOTICE: All authorized recipients of any personal data, personal information, privileged and sensitive personal information contained in this document including other pertinent documents attached thereto that are shared by the Commission on Elections in compliance with the existing laws and rules, and in conformity with the Data Privacy Act of 2012 ( R.A. -

“Slice-Of-Life” Customization of Bankruptcy Models: Belarusian Experience and Future Development
PRACE NAUKOWE Uniwersytetu Ekonomicznego we Wrocławiu RESEARCH PAPERS of Wrocław University of Economics Nr 381 Financial Investments and Insurance – Global Trends and the Polish Market edited by Krzysztof Jajuga Wanda Ronka-Chmielowiec Publishing House of Wrocław University of Economics Wrocław 2015 Copy-editing: Agnieszka Flasińska Layout: Barbara Łopusiewicz Proof-reading: Barbara Cibis Typesetting: Małgorzata Czupryńska Cover design: Beata Dębska Information on submitting and reviewing papers is available on the Publishing House’s website www.pracenaukowe.ue.wroc.pl www.wydawnictwo.ue.wroc.pl The publication is distributed under the Creative Commons Attribution 3.0 Attribution-NonCommercial-NoDerivs CC BY-NC-ND © Copyright by Wrocław University of Economics Wrocław 2015 ISSN 1899-3192 e-ISSN 2392-0041 ISBN 978-83-7695-463-9 The original version: printed Publication may be ordered in Publishing House tel./fax 71 36-80-602; e-mail: [email protected] www.ksiegarnia.ue.wroc.pl Printing: TOTEM Contents Introduction ..................................................................................................... 9 Roman Asyngier: The effect of reverse stock split on the Warsaw Stock Ex- change ......................................................................................................... 11 Monika Banaszewska: Foreign investors on the Polish Treasury bond market in the years 2007-2013 ................................................................................ 26 Katarzyna Byrka-Kita, Mateusz Czerwiński: Large block trades -

Bio::Graphics HOWTO Lincoln Stein Cold Spring Harbor Laboratory1 [email protected]
Bio::Graphics HOWTO Lincoln Stein Cold Spring Harbor Laboratory1 [email protected] This HOWTO describes how to render sequence data graphically in a horizontal map. It applies to a variety of situations ranging from rendering the feature table of a GenBank entry, to graphing the positions and scores of a BLAST search, to rendering a clone map. It describes the programmatic interface to the Bio::Graphics module, and discusses how to create dynamic web pages using Bio::DB::GFF and the gbrowse package. Table of Contents Introduction ...........................................................................................................................3 Preliminaries..........................................................................................................................3 Getting Started ......................................................................................................................3 Adding a Scale to the Image ...............................................................................................5 Improving the Image............................................................................................................7 Parsing Real BLAST Output...............................................................................................8 Rendering Features from a GenBank or EMBL File ....................................................12 A Better Version of the Feature Renderer ......................................................................14 Summary...............................................................................................................................18