A Cephalopod Approach to Rethinking About the Importance of the Bohr and Haldane Effects·
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Would the Hemoglobin–Oxygen Dissociation Curve Shift with Fetal Hemoglobin Compared with Adult Hemoglobin?
❖ CASE 19 A 23-year-old man with no medical problems is brought to the emergency cen- ter by family members who found him to be confused, nauseated, short of breath, and complaining of a headache. The patient was found in the basement of their home next to a furnace, where he was trying to stay warm on a cold winter day. On examination, the patient is lethargic and confused. His lips appear a bright pink. A urine drug screen is obtained and is negative. His serum carboxyhemoglobin level is elevated. The patient is diagnosed with car- bon monoxide poisoning and is admitted to the hospital for further treatment. ◆ What is the mechanism by which carbon monoxide causes hypoxia? ◆ In which direction (right or left) would the hemoglobin–oxygen dissociation curve shift with fetal hemoglobin compared with adult hemoglobin? ◆ What is the most common way in which carbon dioxide is transported in venous blood? 154 CASE FILES: PHYSIOLOGY ANSWERS TO CASE 19: OXYGEN-CARBON DIOXIDE TRANSPORT Summary: A 23-year-old man has confusion, nausea, shortness of breath, and headache after being found near a furnace in the basement. The patient has clinical and laboratory findings consistent with carbon monoxide poisoning. ◆ Hypoxia with carbon monoxide: Decreased oxygen-binding capacity of hemoglobin. ◆ Fetal hemoglobin–oxygen dissociation curve: Shift to the left. ◆ Most common way carbon dioxide is transported in the blood: - Bicarbonate (HCO3 ). CLINICAL CORRELATION Carbon monoxide is a gas that is produced commonly by internal combustion engines, fossil-fuel home appliances (heaters, stoves, furnaces), and incom- plete combustion of nearly all natural and synthetic products. -
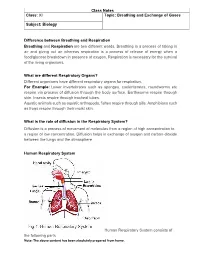
Class Notes Class: XI Topic: Breathing and Exchange of Gases Subject: Biology Difference Between Breathing and Respiration Breat
Class Notes Class: XI Topic: Breathing and Exchange of Gases Subject: Biology Difference between Breathing and Respiration Breathing and Respiration are two different words. Breathing is a process of taking in air and giving out air whereas respiration is a process of release of energy when a food/glucose breakdown in presence of oxygen. Respiration is necessary for the survival of the living organisms. What are different Respiratory Organs? Different organisms have different respiratory organs for respiration. For Example: Lower invertebrates such as sponges, coelenterates, roundworms etc respire via process of diffusion through the body surface. Earthworms respire through skin. Insects respire through tracheal tubes. Aquatic animals such as aquatic arthropods, fishes respire through gills. Amphibians such as frogs respire through their moist skin. What is the role of diffusion in the Respiratory System? Diffusion is a process of movement of molecules from a region of high concentration to a region of low concentration. Diffusion helps in exchange of oxygen and carbon-dioxide between the lungs and the atmosphere. Human Respiratory System Human Respiratory System consists of the following parts Note: The above content has been absolutely prepared from home. • Nostrils is the first part of the respiratory system from where the air enters. • Trachea also known as wind pipe filters the air that is inhaled. Trachea is covered by a lid known as glottis at the time of swallowing of food. It prevents the food from entering into the trachea. It branches into bronchi. • Bronchi also known as air tubes pass air into the lungs. Bronchi branches into bronchioles. -

Respiratory System True Or False
Anatomy & Physiology Test November 4, 2017 Respiratory System True or False: 1. The major factor that stimulates the medulla oblongata and pons to produce respiration is not oxygen concentration, but rather the concentration of carbon dioxide in the blood. 2. The functional residual capacity (FRC) is the amount of air that remains in the lung after a normal tidal expiration; it is the sum of expiratory reserve volume and residual volume. 3. Compared to an atmospheric air, the alveolar air contains less of water vapor but greater amount of carbon dioxide. 4. Binding of the first oxygen molecule causes a conformational change in hemoglobin that allows the second molecule of oxygen to bind more readily, however at the tissue level, as the first oxygen molecule dissociates, it is harder for the next oxygen molecule to dissociate. 5. A large fraction—about 70 percent—of the carbon dioxide molecules that diffuse into the blood is transported to the lungs as dissolved state in plasma. 6. It is more difficult for a body to achieve the same level of oxygen saturation at high altitude than at low altitude, due to lower portion of oxygen (5%). Short Answers: 7. Describe how the rise of concentration of carbon dioxide or hydrogen ions affects the respiration centers and the rate and depth of respiration. 8. Explain what happens to perfusion when ventilation to an alveolus is not sufficient. 9. What are the factors that may cause the oxygen–hemoglobin dissociation curve shift to the “RIGHT”? 10. Please complete the following chemical equation: _____ ______ + - Enzyme:______ 11. -

CO2 Transport Linda Costanzo, Ph.D
CO2 Transport Linda Costanzo, Ph.D. OBJECTIVES: After studying this lecture, the student should understand: 1. How carbon dioxide is carried in blood, especially as bicarbonate. 2. The effect of oxygen on the carbon dioxide content of blood. CO2 is carried in the blood in three forms: dissolved CO2, CO2 bound to proteins such as - hemoglobin (called carbaminohemoglobin) and, most importantly, as HCO3 . I. DISSOLVED CO2 is described by Henry’s law as the partial pressure times the solubility and accounts for 5% of the total CO2 content of blood. The solubility of CO2 in blood is 0.07 ml CO2/100 ml blood/mm Hg (more than twenty times the solubility of O2). Thus, in arterial blood with a PCO2 of 40 mm Hg, dissolved CO2 is: Dissolved CO2 = PCO2 x solubility = 40 mm Hg x 0.07 ml CO2/100 ml blood/mm Hg = 2.8 ml CO2/100 ml blood, or 2.8 vol% II. CARBAMINOHEMOGLOBIN CO2 binds to terminal amino groups on hemoglobin and plasma proteins such as albumin, so-called carbamino compounds. Carbamino compounds account for 3% of the total CO2 in blood, 2/3 of which is carbaminohemoglobin. - III. HCO3 - - 92%, of the CO2 is carried in blood as HCO3 . The reactions that produce HCO3 are as follows: + - CO2 + H2O W H2CO3 W H + HCO3 Carbonic anhydrase In the tissues, CO2 generated from aerobic metabolism is added to venous blood. In the red cells of venous blood, the above reactions occur, generating H+ and - + HCO3 . The H remains inside the red cells, buffered by deoxyhemoglobin. -

A Carbon Monoxide Transport Model of the Human Respiratory System Applied to …
A Carbon Monoxide Transport Model of The Human Respiratory System Applied to … A Carbon Monoxide Transport Model Cyro Albuquerque Neto of the Human Respiratory System [email protected] Applied to Urban Atmosphere Jurandir Itizo Yanagihara Senior Member, ABCM Exposure Analysis [email protected] The aim of this work is to analyze the carbon monoxide (CO) transport in the human body submitted to several physical activity levels. A complete mathematical model of the human respiratory system was developed, considering the exchanges of CO, oxygen (O2) and Fábio Turri carbon dioxide (CO2) in the lung, blood and tissues. The human body was divided in the [email protected] following compartments: alveolar, pulmonary capillaries, arterial, venous, tissue capillary University of Sao Paulo - USP and tissues. The gas transport in the blood and tissues is represented by empirical Escola Politécnica equations. The physiological parameters were described in function of physical activity. Department of Mechanical Engineering The model was validated by comparing its results with experimental data of controlled CO 055809-900 São Paulo, SP, Brazil exposition. The agreement was excellent. CO concentration curves for critical days of São Caetano do Sul city (SP, Brazil) atmosphere were used as model input. The simulation results for some physical activities show that the more intense the activity, the larger the blood carboxyhemoglobin (COHb) level variations. The COHb level was compared with a CO quality air criteria, which showed to be adequate for low and moderate physical activity levels. Keywords: compartment model, carbon monoxide, gas transport, respiratory system, air pollution The models of Selvakumar, Sharan and Singh (1992) and Introduction Sharan and Selvakumar (1999) divide the lung into compartments, one alveolar and several pulmonary capillaries. -

Respiratory System.Pdf
Respiratory System Respiratory System - Overview: Assists in the detection Protects system of odorants Respiratory (debris / pathogens / dessication) System 5 3 4 Produces sound (vocalization) Provides surface area for gas exchange (between air / blood) 1 2 For the body to survive, there must be a constant supply of Moves air to / from gas O2 and a constant exchange surface disposal of CO 2 Marieb & Hoehn (Human Anatomy and Physiology, 8th ed.) – Table 19.1 Respiratory System Respiratory System Functional Anatomy: Functional Anatomy: Trachea Epiglottis Naming of pathways: • > 1 mm diameter = bronchus Upper Respiratory • Conduction of air • < 1 mm diameter = bronchiole System • Gas exchange Primary • < 0.5 mm diameter = terminal bronchiole Bronchus • Filters / warms / humidifies Lower Respiratory Bronchi System incoming air bifurcation (23 orders) 1) External nares 5) Larynx 2) Nasal cavity • Provide open airway Green = Conducting zone • Resonance chamber • channel air / food Purple = Respiratory zone 3) Uvula • voice production (link) 4) Pharynx 6) Trachea 7) Bronchial tree • Nasopharynx Bronchiole 8) Alveoli • Oropharynx Terminal Bronchiole Respiratory Bronchiole • Laryngopharynx Alveolus Martini et. al. (Fundamentals of Anatomy and Physiology, 7th ed.) – Figure 23.1 Martini et. al. (Fundamentals of Anatomy and Physiology, 7th ed.) – Figure 23.9 Respiratory System Respiratory System Functional Anatomy: Functional Anatomy: Respiratory Mucosa / Submucosa: How are inhaled debris / pathogens cleared from respiratory tract? Near Near trachea alveoli Nasal Cavity: Epithelium: Particles > 10 µm Pseudostratified Simple columnar cuboidal Conducting Zone: Particles 5 – 10 µm Cilia No cilia Respiratory Zone: Mucus Escalator Particles 1 – 5 µm Mucosa: Lamina Propria (areolar tissue layer): Mucous membrane (epithelium / areolar tissue) smooth smooth muscle muscle Mucous No glands mucous glands Cartilage: Rings Plates / none Macrophages Martini et. -

Hypercapnia from Hyperoxia in COPD: Another Piece of the Puzzle Or Another Puzzle Entirely?
Hypercapnia From Hyperoxia in COPD: Another Piece of the Puzzle or Another Puzzle Entirely? 4 ˙ Embarrassment is a powerful teacher. I was barely et al found that the reduction in minute ventilation (VE) 3 months into my internship, on night float at my residen- was transient and inadequate to explain the degree of hy- cy’s Veterans Affairs hospital, when I received a page percapnia. from a concerned ward nurse. A patient was unresponsive. In another study, Aubier et al5 found no correlation be- ˙ He was admitted for a severe COPD exacerbation. I ex- tween the degree of hypercapnia and the decrease in VE. amined him, and he only groaned with a vigorous sternal Instead, they hypothesized that the excess hypercapnia was rub. Suspecting hypercapnic narcosis, I drew an arterial caused by 2 factors. The first was the Haldane effect (ie, blood gas. The results confirmed my suspicion. The pa- the release of CO2 when deoxyhemoglobin converts to tient was suffering from acute respiratory acidosis. In a panic, I paged the resident taking ICU admissions. The patient needed to go to the ICU and be intubated. The SEE THE ORIGINAL STUDY ON PAGE 328 resident said he would see the patient. He wordlessly slipped into the room and went straight to the oxygen flow meter. He turned the flow down several liters and waited. He did oxyhemoglobin). This mechanism was later confirmed by not have to wait long: it was as if the patient had received Luft et al.6 The second was a decrease in pulmonary ven- a naloxone injection. -

Diffusion and Transport of Respiratory Gases
SEMESTER – IV ZOO CC409 : Animal Physiology: Life Sustaining Systems, Unit 2 Diffusion and Transport of Respiratory Gases Prof (Dr.) Shahla Yasmin Department of Zoology Patna Women’s College Patna University Learning objectives The students will learn: • Structure of respiratory membrane • Characteristics of respiratory membrane • Diffusion and transport of oxygen • Diffusion and transport of carbon-di-oxide • Chloride shift mechanism • Haldane effect • Diffusion of respiratory gases occurs through the respiratory membrane. • This “air-blood barrier” (the respiratory membrane) is where gas exchange occurs via passive diffusion – oxyygen diffuses from air in alveolus (singular of alveoli) to blood in capillary – Carbon dioxide diffuses from the blood in the capillary into the air in the alveolus - Fig: Respiratory Gas exchange area 3 Epithelial basement Capillary basement membrane Interstitial membrane space Capillary endothelium Alveolar epithelium Fluid and Red surfactant blood layer cell Alveolus Capillary Diffusion O2 Diffusion CO2 Structure of respiratory membrane Features of Respiratory membrane • Thickness = 0.6um (15 X thinner than tissue paper) • Air filled alveoli account for most of the lung volume • Very great area for gas exchange =50-100 sqm • Quantity of blood in the lung capillaries=60-140ml Respiratory bronchiole Alveolar duct Alveoli Alveolar sac 5 Features of Respiratory membrane • Average diameter of pulmonary capillary=5um • RBC size=7.5um (can squeeze through pulmonary capillary) • Respiratory gases are highly soluble in lipids and thus in cell membrane and thus can easily pass through respiratory membrane • All the above mentioned features facilitate rapid gaseous exchange across the respiratory membrane Factors that affect the rate of gas diffusion through the respiratory membrane: 1. -

Pulmonary Ventilation
CHAPTER 2 Pulmonary Ventilation © IT Stock/Polka Dot/ inkstock Chapter Objectives By studying this chapter, you should be able to do Name two things that cause pleural pressure the following: to decrease. 8. Describe the mechanics of ventilation with 1. Identify the basic structures of the conducting respect to the changes in pulmonary pressures. and respiratory zones of the ventilation system. 9. Identify the muscles involved in inspiration and 2. Explain the role of minute ventilation and its expiration at rest. relationship to the function of the heart in the 10. Describe the partial pressures of oxygen and production of energy at the tissues. carbon dioxide in the alveoli, lung capillaries, 3. Identify the diff erent ways in which carbon tissue capillaries, and tissues. dioxide is transported from the tissues to the 11. Describe how carbon dioxide is transported in lungs. the blood. 4. Explain the respiratory advantage of breathing 12. Explain the significance of the oxygen– depth versus rate during a treadmill exercise. hemoglobin dissociation curve. 5. Describe the composition of ambient air and 13. Discuss the eff ects of decreasing pH, increasing alveolar air and the pressure changes in the pleu- temperature, and increasing 2,3-diphosphoglyc- ral and pulmonary spaces. erate on the HbO dissociation curve. 6. Diagram the three ways in which carbon dioxide 2 14. Distinguish between and explain external res- is transported in the venous blood to the lungs. piration and internal respiration. 7. Defi ne pleural pressure. What happens to alve- olar volume when pleural pressure decreases? Chapter Outline Pulmonary Structure and Function Pulmonary Volumes and Capacities Anatomy of Ventilation Lung Volumes and Capacities Lungs Pulmonary Ventilation Mechanics of Ventilation Minute Ventilation Inspiration Alveolar Ventilation Expiration Pressure Changes Copyright ©2014 Jones & Bartlett Learning, LLC, an Ascend Learning Company Content not final. -

Diving Physiology 3
Diving Physiology 3 SECTION PAGE SECTION PAGE 3.0 GENERAL ...................................................3- 1 3.3.3.3 Oxygen Toxicity ........................3-21 3.1 SYSTEMS OF THE BODY ...............................3- 1 3.3.3.3.1 CNS: Central 3.1.1 Musculoskeletal System ............................3- 1 Nervous System .........................3-21 3.1.2 Nervous System ......................................3- 1 3.3.3.3.2 Lung and 3.1.3 Digestive System.....................................3- 2 “Whole Body” ..........................3-21 3.2 RESPIRATION AND CIRCULATION ...............3- 2 3.2.1 Process of Respiration ..............................3- 2 3.3.3.3.3 Variations In 3.2.2 Mechanics of Respiration ..........................3- 3 Tolerance .................................3-22 3.2.3 Control of Respiration..............................3- 4 3.3.3.3.4 Benefits of 3.2.4 Circulation ............................................3- 4 Intermittent Exposure..................3-22 3.2.4.1 Blood Transport of Oxygen 3.3.3.3.5 Concepts of and Carbon Dioxide ......................3- 5 Oxygen Exposure 3.2.4.2 Tissue Gas Exchange.....................3- 6 Management .............................3-22 3.2.4.3 Tissue Use of Oxygen ....................3- 6 3.3.3.3.6 Prevention of 3.2.5 Summary of Respiration CNS Poisoning ..........................3-22 and Circulation Processes .........................3- 8 3.2.6 Respiratory Problems ...............................3- 8 3.3.3.3.7 The “Oxygen Clock” 3.2.6.1 Hypoxia .....................................3- -

Acid-Base Disturbances in Patients with Asthma: a Literature Review and Comments on Their Pathophysiology
Journal of Clinical Medicine Review Acid-Base Disturbances in Patients with Asthma: A Literature Review and Comments on Their Pathophysiology Ioannis Vasileiadis 1,*, Emmanouil Alevrakis 2, Sevasti Ampelioti 3, Dimitrios Vagionas 1, Nikoletta Rovina 1 and Antonia Koutsoukou 1 1 Intensive Care Unit, 1st Department of Respiratory Medicine, National and Kapodistrian University of Athens, Sotiria Hospital, 115 27 Athens, Greece; [email protected] (D.V.); [email protected] (N.R.); [email protected] (A.K.) 2 4th Department of Respiratory Medicine, Sotiria Hospital, 115 27 Athens, Greece; [email protected] 3 5th Department of Respiratory Medicine, Sotiria Hospital, 115 27 Athens, Greece; [email protected] * Correspondence: [email protected] or [email protected]; Tel: +30-697-764-4866 or +30-210-7763725 ; Fax: +30-210-778-1250 Received: 27 March 2019; Accepted: 23 April 2019; Published: 25 April 2019 Abstract: Asthma is a common illness throughout the world that affects the respiratory system function, i.e., a system whose operational adequacy determines the respiratory gases exchange. It is therefore expected that acute severe asthma will be associated with respiratory acid-base disorders. In addition, the resulting hypoxemia along with the circulatory compromise due to heart–lung interactions can reduce tissue oxygenation, with a particular impact on respiratory muscles that have increased energy needs due to the increased workload. Thus, anaerobic metabolism may ensue, leading to lactic acidosis. Additionally, chronic hypocapnia in asthma can cause a compensatory drop in plasma bicarbonate concentration, resulting in non-anion gap acidosis. Indeed, studies have shown that in acute severe asthma, metabolic acid-base disorders may occur, i.e., high anion gap or non-anion gap metabolic acidosis. -

Gas Exchange and Ventilation–Perfusion Relationships in the Lung
ERJ Express. Published on July 25, 2014 as doi: 10.1183/09031936.00037014 REVIEW IN PRESS | CORRECTED PROOF Gas exchange and ventilation–perfusion relationships in the lung Johan Petersson1,2 and Robb W. Glenny3,4 Affiliations: 1Section of Anaesthesiology and Intensive Care Medicine, Dept of Physiology and Pharmacology, Karolinska Institutet, Stockholm, Sweden. 2Dept of Anaesthesiology, Surgical Services and Intensive Care, Karolinska University Hospital, Stockholm, Sweden. 3Dept of Medicine, Division of Pulmonary and Critical Care Medicine, University of Washington, Seattle, WA, USA. 4Dept of Physiology and Biophysics, University of Washington, Seattle, WA, USA. Correspondence: Johan Petersson, Dept of Anaesthesiology, Surgical Services and Intensive Care, Karolinska University Hospital, 17176 Stockholm, Sweden. E-mail: [email protected] ABSTRACT This review provides an overview of the relationship between ventilation/perfusion ratios and gas exchange in the lung, emphasising basic concepts and relating them to clinical scenarios. For each gas exchanging unit, the alveolar and effluent blood partial pressures of oxygen and carbon dioxide (PO2 and PCO2) are determined by the ratio of alveolar ventilation to blood flow (V9A/Q9) for each unit. Shunt and low V9A/Q9 regions are two examples of V9A/Q9 mismatch and are the most frequent causes of hypoxaemia. Diffusion limitation, hypoventilation and low inspired PO2 cause hypoxaemia, even in the absence of V9A/Q9 mismatch. In contrast to other causes, hypoxaemia due to shunt responds poorly to supplemental oxygen. Gas exchanging units with little or no blood flow (high V9A/Q9 regions) result in alveolar dead space and increased wasted ventilation, i.e. less efficient carbon dioxide removal.