Nerine Bowdenii
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

Summary of Offerings in the PBS Bulb Exchange, Dec 2012- Nov 2019
Summary of offerings in the PBS Bulb Exchange, Dec 2012- Nov 2019 3841 Number of items in BX 301 thru BX 463 1815 Number of unique text strings used as taxa 990 Taxa offered as bulbs 1056 Taxa offered as seeds 308 Number of genera This does not include the SXs. Top 20 Most Oft Listed: BULBS Times listed SEEDS Times listed Oxalis obtusa 53 Zephyranthes primulina 20 Oxalis flava 36 Rhodophiala bifida 14 Oxalis hirta 25 Habranthus tubispathus 13 Oxalis bowiei 22 Moraea villosa 13 Ferraria crispa 20 Veltheimia bracteata 13 Oxalis sp. 20 Clivia miniata 12 Oxalis purpurea 18 Zephyranthes drummondii 12 Lachenalia mutabilis 17 Zephyranthes reginae 11 Moraea sp. 17 Amaryllis belladonna 10 Amaryllis belladonna 14 Calochortus venustus 10 Oxalis luteola 14 Zephyranthes fosteri 10 Albuca sp. 13 Calochortus luteus 9 Moraea villosa 13 Crinum bulbispermum 9 Oxalis caprina 13 Habranthus robustus 9 Oxalis imbricata 12 Haemanthus albiflos 9 Oxalis namaquana 12 Nerine bowdenii 9 Oxalis engleriana 11 Cyclamen graecum 8 Oxalis melanosticta 'Ken Aslet'11 Fritillaria affinis 8 Moraea ciliata 10 Habranthus brachyandrus 8 Oxalis commutata 10 Zephyranthes 'Pink Beauty' 8 Summary of offerings in the PBS Bulb Exchange, Dec 2012- Nov 2019 Most taxa specify to species level. 34 taxa were listed as Genus sp. for bulbs 23 taxa were listed as Genus sp. for seeds 141 taxa were listed with quoted 'Variety' Top 20 Most often listed Genera BULBS SEEDS Genus N items BXs Genus N items BXs Oxalis 450 64 Zephyranthes 202 35 Lachenalia 125 47 Calochortus 94 15 Moraea 99 31 Moraea -

Boophone Disticha
Micropropagation and pharmacological evaluation of Boophone disticha Lee Cheesman Submitted in fulfilment of the academic requirements for the degree of Doctor of Philosophy Research Centre for Plant Growth and Development School of Life Sciences University of KwaZulu-Natal, Pietermaritzburg April 2013 COLLEGE OF AGRICULTURE, ENGINEERING AND SCIENCES DECLARATION 1 – PLAGIARISM I, LEE CHEESMAN Student Number: 203502173 declare that: 1. The research contained in this thesis, except where otherwise indicated, is my original research. 2. This thesis has not been submitted for any degree or examination at any other University. 3. This thesis does not contain other persons’ data, pictures, graphs or other information, unless specifically acknowledged as being sourced from other persons. 4. This thesis does not contain other persons’ writing, unless specifically acknowledged as being sourced from other researchers. Where other written sources have been quoted, then: a. Their words have been re-written but the general information attributed to them has been referenced. b. Where their exact words have been used, then their writing has been placed in italics and inside quotation marks, and referenced. 5. This thesis does not contain text, graphics or tables copied and pasted from the internet, unless specifically acknowledged, and the source being detailed in the thesis and in the reference section. Signed at………………………………....on the.....….. day of ……......……….2013 ______________________________ SIGNATURE i STUDENT DECLARATION Micropropagation and pharmacological evaluation of Boophone disticha I, LEE CHEESMAN Student Number: 203502173 declare that: 1. The research reported in this dissertation, except where otherwise indicated is the result of my own endeavours in the Research Centre for Plant Growth and Development, School of Life Sciences, University of KwaZulu-Natal, Pietermaritzburg. -

Guide to the Flora of the Carolinas, Virginia, and Georgia, Working Draft of 17 March 2004 -- LILIACEAE
Guide to the Flora of the Carolinas, Virginia, and Georgia, Working Draft of 17 March 2004 -- LILIACEAE LILIACEAE de Jussieu 1789 (Lily Family) (also see AGAVACEAE, ALLIACEAE, ALSTROEMERIACEAE, AMARYLLIDACEAE, ASPARAGACEAE, COLCHICACEAE, HEMEROCALLIDACEAE, HOSTACEAE, HYACINTHACEAE, HYPOXIDACEAE, MELANTHIACEAE, NARTHECIACEAE, RUSCACEAE, SMILACACEAE, THEMIDACEAE, TOFIELDIACEAE) As here interpreted narrowly, the Liliaceae constitutes about 11 genera and 550 species, of the Northern Hemisphere. There has been much recent investigation and re-interpretation of evidence regarding the upper-level taxonomy of the Liliales, with strong suggestions that the broad Liliaceae recognized by Cronquist (1981) is artificial and polyphyletic. Cronquist (1993) himself concurs, at least to a degree: "we still await a comprehensive reorganization of the lilies into several families more comparable to other recognized families of angiosperms." Dahlgren & Clifford (1982) and Dahlgren, Clifford, & Yeo (1985) synthesized an early phase in the modern revolution of monocot taxonomy. Since then, additional research, especially molecular (Duvall et al. 1993, Chase et al. 1993, Bogler & Simpson 1995, and many others), has strongly validated the general lines (and many details) of Dahlgren's arrangement. The most recent synthesis (Kubitzki 1998a) is followed as the basis for familial and generic taxonomy of the lilies and their relatives (see summary below). References: Angiosperm Phylogeny Group (1998, 2003); Tamura in Kubitzki (1998a). Our “liliaceous” genera (members of orders placed in the Lilianae) are therefore divided as shown below, largely following Kubitzki (1998a) and some more recent molecular analyses. ALISMATALES TOFIELDIACEAE: Pleea, Tofieldia. LILIALES ALSTROEMERIACEAE: Alstroemeria COLCHICACEAE: Colchicum, Uvularia. LILIACEAE: Clintonia, Erythronium, Lilium, Medeola, Prosartes, Streptopus, Tricyrtis, Tulipa. MELANTHIACEAE: Amianthium, Anticlea, Chamaelirium, Helonias, Melanthium, Schoenocaulon, Stenanthium, Veratrum, Toxicoscordion, Trillium, Xerophyllum, Zigadenus. -

Differences in Floral Development Between Lycoris Radiata And
R ESEARCH ARTICLE doi: 10.2306/scienceasia1513-1874.2020.032 Differences in floral development between Lycoris radiata and Lycoris sprengeri a,† b,† c d a a,e, Junhuo Cai , Junjun Fan , Xuying Wei , Donglin Zhang , Jiajia Ren , Lu Zhang ∗ a College of Landscape and Art, Jiangxi Agricultural University, Nanchang 330045 China b College of Horticulture, Jinling Institute of Technology, Nanjing 210037 China c College of Art, Jiangxi Finance and Economics University, Nanchang 330032 China d Department of Horticulture, University of Georgia, Athens, GA 30602 USA e Collaboration Innovation Center of Jiangxi Typical Trees Cultivation and Utilization, Jiangxi Agricultural University, Nanchang 330045 China ∗Corresponding author, e-mail: [email protected], Cai J. and Fan J. contributed equally to this paper. Received 12 Sep 2019 Accepted 21 Mar 2020 ABSTRACT: Lycoris radiata and Lycoris sprengeri are two typical plants of the genus Lycoris. They have different leafing and flowering patterns. To understand the habits of Lycoris species and regulate the flowering period effectively, we compared flower bud development of these two species using phenological, anatomical, and physiological measurements. Our results showed there were significant differences in phenology between the two species. The sprouting stage of L. radiata took place in autumn, the leaf growth period lasted seven months, dormancy lasted approximately 100 days before flowering, and there was no dormancy period after flowering. In comparison, the sprouting period of L. sprengeri was delayed until the spring of the following year, the leaf growth stage lasted approximately 3 months, the flowering stage took place from Jul–Aug, and dormancy lasted 1–2 months before flowering and for another 6 months after flowering. -

Comparative Chloroplast Genomes of Four Lycoris Species (Amaryllidaceae) Provides New Insight Into Interspecific Relationship and Phylogeny
biology Article Comparative Chloroplast Genomes of Four Lycoris Species (Amaryllidaceae) Provides New Insight into Interspecific Relationship and Phylogeny Fengjiao Zhang 1,2, Ning Wang 1,2, Guanghao Cheng 1,2, Xiaochun Shu 1,2, Tao Wang 1,2 , Weibing Zhuang 1,2, Ruisen Lu 1,2,* and Zhong Wang 1,2,* 1 Jiangsu Key Laboratory for the Research and Utilization of Plant Resources, Institute of Botany, Jiangsu Province and Chinese Academy of Sciences (Nanjing Botanical Garden Mem. Sun Yat-Sen), Nanjing 210014, China; [email protected] (F.Z.); [email protected] (N.W.); [email protected] (G.C.); [email protected] (X.S.); [email protected] (T.W.); [email protected] (W.Z.) 2 The Jiangsu Provincial Platform for Conservation and Utilization of Agricultural Germplasm, Institute of Botany, Jiangsu Province and Chinese Academy of Sciences (Nanjing Botanical Garden Mem. Sun Yat-Sen), Nanjing 210014, China * Correspondence: [email protected] (R.L.); [email protected] (Z.W.) Simple Summary: The genus Lycoris (Amaryllidaceae) comprises about 20 species with high orna- mental and medicinal value. However, germplasm identification is still difficult due to frequent interspecific hybridization and intraspecific morphological variation within this genus. Plastid genome sequencing has been proven to be a useful tool to identify closely related species and is widely used in the field of plant evolution and phylogeny. In the present study, we provided four Citation: Zhang, F.; Wang, N.; chloroplast genomes of Lycoris and retrieved seven published species in the genus for comparative Cheng, G.; Shu, X.; Wang, T.; Zhuang, genomics and phylogenetic analyses. All these chloroplast genomes possess the typical quadripartite W.; Lu, R.; Wang, Z. -

STUDY on GROWTH, DEVELOPMENT and SOME BIOCHEMICAL ASPECTS of SEVERAL VARIETIES of Nerine
• STUDY ON GROWTH, DEVELOPMENT AND SOME BIOCHEMICAL ASPECTS OF SEVERAL VARIETIES OF Nerine by KUMALA DEWI partia,1 Submitted in,fulfilment of the requirements for the A degree of Master of Science Studies Department of Plant Science University of Tasmania May, 1993 DECLARATION To the best of my knowledge and belief, this thesis contains no material which has been submitted for the award of any other degree or diploma, nor does it contain any paraphrase of previously published material except where due reference is made in the text. Kumala Dewi ii ABSTRACT Nerine fothergillii bulbs were stored at different temperatures for a certain period of time and then planted and grown in an open condition. The effect of the different storage temperatures on carbohydrate -; content and endogenous gibberellins w Ct,5 examined in relation to flowering. Flowering percentage and flower number in each umbel was reduced when the bulbs were stored at 300 C while bulbs which received 50 C treatment possess earlier flowering and longer flower stalksthan bulbs without 5 0 C storage treatment. Carbohydrates in both outer and inner scales of N. fothergillii were examined semi-quantitatively by paper chromatography. Glucose, fructose and sucrose have been identified from paper chromatogramS. Endogenous gibberellins in N. fothergillii have been identified by GC - SIM and full mass spectra from GCMS. These include GA19, GA20 and G Al, their presence suggests the occurence of the early 13 - hydroxylation pathway. The response of N. bowdenii grown under Long Day (LD) and Short Day (SD) conditions w as studied. Ten plants from each treatment were examined at intervalsof 4 weeks. -

Complete Chloroplast Genomes Shed Light on Phylogenetic
www.nature.com/scientificreports OPEN Complete chloroplast genomes shed light on phylogenetic relationships, divergence time, and biogeography of Allioideae (Amaryllidaceae) Ju Namgung1,4, Hoang Dang Khoa Do1,2,4, Changkyun Kim1, Hyeok Jae Choi3 & Joo‑Hwan Kim1* Allioideae includes economically important bulb crops such as garlic, onion, leeks, and some ornamental plants in Amaryllidaceae. Here, we reported the complete chloroplast genome (cpDNA) sequences of 17 species of Allioideae, fve of Amaryllidoideae, and one of Agapanthoideae. These cpDNA sequences represent 80 protein‑coding, 30 tRNA, and four rRNA genes, and range from 151,808 to 159,998 bp in length. Loss and pseudogenization of multiple genes (i.e., rps2, infA, and rpl22) appear to have occurred multiple times during the evolution of Alloideae. Additionally, eight mutation hotspots, including rps15-ycf1, rps16-trnQ-UUG, petG-trnW-CCA , psbA upstream, rpl32- trnL-UAG , ycf1, rpl22, matK, and ndhF, were identifed in the studied Allium species. Additionally, we present the frst phylogenomic analysis among the four tribes of Allioideae based on 74 cpDNA coding regions of 21 species of Allioideae, fve species of Amaryllidoideae, one species of Agapanthoideae, and fve species representing selected members of Asparagales. Our molecular phylogenomic results strongly support the monophyly of Allioideae, which is sister to Amaryllioideae. Within Allioideae, Tulbaghieae was sister to Gilliesieae‑Leucocoryneae whereas Allieae was sister to the clade of Tulbaghieae‑ Gilliesieae‑Leucocoryneae. Molecular dating analyses revealed the crown age of Allioideae in the Eocene (40.1 mya) followed by diferentiation of Allieae in the early Miocene (21.3 mya). The split of Gilliesieae from Leucocoryneae was estimated at 16.5 mya. -

Sell Cut Flowers from Perennial Summer-Flowering Bulbs
SELL CUT FLOWERS FROM PERENNIAL SUMMER-FLOWERING BULBS Andy Hankins Extension Specialist-Alternative Agriculture, Virginia State University Reviewed by Chris Mullins, Virginia State University 2018 Commercial producers of field-grown flower cut flowers generally have a wide selection of crops to sell in April, May and June. Many species of annual and especially perennial cut flowers bloom during these three months. Many flower crops are sensitive to day length. Crops that bloom during long days such as larkspur, yarrow, peonies and gypsophila cannot be made to bloom after the summer equinox on June 21st. Other crops such as snapdragons may be day length neutral but they are adversely affected by the very warm days and nights of mid-summer. It is much more challenging for Virginia cut flower growers to have a diverse selection of flower crops for marketing from July to September when day length is getting shorter and day temperatures are getting hotter. Quite a few growers offer the same inventory of sunflowers, zinnias, celosia and gladiolas during the middle of the summer because everything else has come and gone. A group of plants that may offer new opportunities for sales of cut flowers during mid-summer are summer-flowering bulbs. Many of these summer-flowering bulbs are tropical plants that have only become available in the United States during the last few years. The first question that growers should ask about any tropical plant recommended for field planting is, " Will this species be winter hardy in Virginia?" Many of the bulb species described in this article are not very winter hardy. -
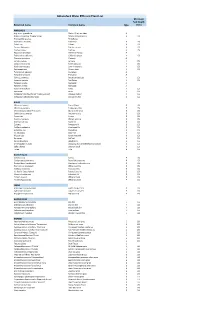
Abbotsford Water Efficient Plant List Minimum Soil Depth Botanical Name Common Name Type (Mm)
Abbotsford Water Efficient Plant List Minimum Soil Depth Botanical name Common name type (mm) ANNUALS Argemone grandifolia Statice/Sea Lavendar A Begonia x hybrida 'Dragon Wings' Dragon Wing begonia A 150 Bracteantha species Strawflower Calendula officinalis Calendula A 150 Coleus ssp. Coleus A 150 Cosmos bipinnatus Garden cosmos A 150 Cuphea llavea Cuphea A 150 Dyssocua tenuiloba Dahlberg Daisey A Eschscholzia californica California poppy A 150 Gazania spendens Gazania A Lantana camara Lantana A 150 Lobularia maritima Sweet alyssum A 150 Nigella damascena Love-in-the-mist A 150 Oesteopermum African daisy A 150 Pelargonium species Geranium A Portulaca oleracea Portulaca A Salvia guaranitica Anise-scented sage A 150 Scaevola aemula Fan flower A 150 Targetes erecta Marogold A Targetes erecta Marogold A Viola x wittrockiana Pansy A 150 Zinnia ssp. Zinnia A 150 Verbascum bombyciferum 'Arctic Summer' Broussa mullein A 150 Verbascum phlomoides 'Spica' Orange mullein A 150 BULBS Allium christophii Star of Persia B 150 Allium karataviense Turkestan onion B 150 Chionodoxa forbesii 'Pink Giant' Glory of the snow B 150 Colchicum autumnale Autumn crocus B 150 Crocus ssp. Crocus B 150 Eranthis hyemalis Winter aconite B 150 Erythronium ssp. Fawn lily B 150 Eucomis Pineapple lily B 150 Fritillaria meleagris Checkered lily B 150 Galanthus ssp. Snowdrop B 150 Iris reticulata Dwarf iris B 150 Muscari ssp. Grape hyacinth B 150 Narcissus Daffodil B 150 Nerine bowdenii Bowden lily B 150 Ornithogalum nutans Drooping Star of Bethlehem/Silverbells B 150 Scilla -

A Molecular Phylogeny of the Genus Scadoxus Raf. (Amaryllidaceae)
Fireball lilies of Africa: a molecular phylogeny of the genus Scadoxus Raf. (Amaryllidaceae) Kine Hals Bødker Master of Science Thesis Natural History Museum, University of Oslo Centre for Ecological and Evolutionary Synthesis, Department of Biosciences, University of Oslo June 15th 2020 © Kine Hals Bødker 2020 Fireball lilies of Africa: a molecular phylogeny of the genus Scadoxus Raf. (Amaryllidaceae) Kine Hals Bødker http://www.duo.uio.no/ Print: Reprosentralen, University of Oslo II Illustration: Aasne Aarhus, 1976. III IV Acknowledgements It has been an exciting journey working with the most beautiful plant genus of this world. There are many people I would like to thank for being a part of this journey. First and foremost, I want to thank my wonderful supervisors for all their help and support over the past two years. I could not have had a better team. My main supervisor, Charlotte - for your enthusiasm, knowledge, support and the most amazing field trips. In the 1970’s, before I was even born, Inger (and colleagues) worked with Scadoxus, which set the stage for this master thesis. I would like to thank Inger for being my additional supervisor and ultimately giving me this opportunity, and helping me understand more of Scadoxus morphology. Anne – for helping with the analyses, and especially for the incredibly helpful checking of spelling, grammatical errors and also helping me discourse my occasionally overwhelming results. Prof. Clemence Zimudzi and Dr. Tesfaye Awas – for fantastic field work experiences in Zimbabwe and Ethiopia, respectively. I would also like to thank the little kids in Ethiopia who helped us find Scadoxus specimens in places we never would have found without them. -

JUDD W.S. Et. Al. (1999) Plant Systematics
CHAPTER8 Phylogenetic Relationships of Angiosperms he angiosperms (or flowering plants) are the dominant group of land Tplants. The monophyly of this group is strongly supported, as dis- cussed in the previous chapter, and these plants are possibly sister (among extant seed plants) to the gnetopsids (Chase et al. 1993; Crane 1985; Donoghue and Doyle 1989; Doyle 1996; Doyle et al. 1994). The angio- sperms have a long fossil record, going back to the upper Jurassic and increasing in abundance as one moves through the Cretaceous (Beck 1973; Sun et al. 1998). The group probably originated during the Jurassic, more than 140 million years ago. Cladistic analyses based on morphology, rRNA, rbcL, and atpB sequences do not support the traditional division of angiosperms into monocots (plants with a single cotyledon, radicle aborting early in growth with the root system adventitious, stems with scattered vascular bundles and usually lacking secondary growth, leaves with parallel venation, flow- ers 3-merous, and pollen grains usually monosulcate) and dicots (plants with two cotyledons, radicle not aborting and giving rise to mature root system, stems with vascular bundles in a ring and often showing sec- ondary growth, leaves with a network of veins forming a pinnate to palmate pattern, flowers 4- or 5-merous, and pollen grains predominantly tricolpate or modifications thereof) (Chase et al. 1993; Doyle 1996; Doyle et al. 1994; Donoghue and Doyle 1989). In all published cladistic analyses the “dicots” form a paraphyletic complex, and features such as two cotyle- dons, a persistent radicle, stems with vascular bundles in a ring, secondary growth, and leaves with net venation are plesiomorphic within angio- sperms; that is, these features evolved earlier in the phylogenetic history of tracheophytes. -

Introducing Lycoris to U.S. Flower Lovers
PEGGY GREB (D300-1) Introducing Lycoris to Golden-yellow fl owers of Lycoris aurea. U.S. Flower Lovers or more than 20 years ARS horticulturist Mark Roh has been intrigued by the origins and habitats of the exotic and beautiful Lycoris. Though various Lycoris species have been grown as ornamentals in China, F Korea, and Japan for many centuries, only two species are readily available here: L. squamigera and L. radiata. They—and the rarer L. incarnata, L. chejuensis, and L. flavescens—are maintained at the U.S. National Arboretum (USNA), in Washington, D.C., and in Beltsville, Maryland. In 1984, Roh collected several unidentified Lycoris species from Anduck Valley, on Korea’s Jeju Island. This subtropical area hosts about 4,000 species of plants. Then in 1998, more Lycoris species were collected in Japan, Korea, and China. DNA molecular markers and chromosome studies proved that some of the unidentified Lycoris collected from Anduck Valley were L. incarnata, a species previously known to be native only to China. It is possible that this accession was brought from China to Korea by bulb collectors, but no record of that can be found. In the past 6 years, extensive visits to areas near Anduck Valley have been made by Roh or by collaborators Mun Seok Seong, of Jeju-do Agricultural Research and Extension Services, and Yong Bong Park, 12 Agricultural Research/December 2005 387664.indd 12 11/11/05 6:23:47 AM PEGGY GREB (D303-1) of Jeju National University. But they have of drought and waterlogging, as well as failed to locate any more L.