STUDY on GROWTH, DEVELOPMENT and SOME BIOCHEMICAL ASPECTS of SEVERAL VARIETIES of Nerine
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Summary of Offerings in the PBS Bulb Exchange, Dec 2012- Nov 2019
Summary of offerings in the PBS Bulb Exchange, Dec 2012- Nov 2019 3841 Number of items in BX 301 thru BX 463 1815 Number of unique text strings used as taxa 990 Taxa offered as bulbs 1056 Taxa offered as seeds 308 Number of genera This does not include the SXs. Top 20 Most Oft Listed: BULBS Times listed SEEDS Times listed Oxalis obtusa 53 Zephyranthes primulina 20 Oxalis flava 36 Rhodophiala bifida 14 Oxalis hirta 25 Habranthus tubispathus 13 Oxalis bowiei 22 Moraea villosa 13 Ferraria crispa 20 Veltheimia bracteata 13 Oxalis sp. 20 Clivia miniata 12 Oxalis purpurea 18 Zephyranthes drummondii 12 Lachenalia mutabilis 17 Zephyranthes reginae 11 Moraea sp. 17 Amaryllis belladonna 10 Amaryllis belladonna 14 Calochortus venustus 10 Oxalis luteola 14 Zephyranthes fosteri 10 Albuca sp. 13 Calochortus luteus 9 Moraea villosa 13 Crinum bulbispermum 9 Oxalis caprina 13 Habranthus robustus 9 Oxalis imbricata 12 Haemanthus albiflos 9 Oxalis namaquana 12 Nerine bowdenii 9 Oxalis engleriana 11 Cyclamen graecum 8 Oxalis melanosticta 'Ken Aslet'11 Fritillaria affinis 8 Moraea ciliata 10 Habranthus brachyandrus 8 Oxalis commutata 10 Zephyranthes 'Pink Beauty' 8 Summary of offerings in the PBS Bulb Exchange, Dec 2012- Nov 2019 Most taxa specify to species level. 34 taxa were listed as Genus sp. for bulbs 23 taxa were listed as Genus sp. for seeds 141 taxa were listed with quoted 'Variety' Top 20 Most often listed Genera BULBS SEEDS Genus N items BXs Genus N items BXs Oxalis 450 64 Zephyranthes 202 35 Lachenalia 125 47 Calochortus 94 15 Moraea 99 31 Moraea -

Influence of Mulching and Foliar Nutrition on The
Journal of Elementology ISSN 1644-2296 Błażewicz-Woźniak M., Wach D., Najda A., Mucha S. 2019. Influence of mulching and foliar nutrition on the formation of bulbs and content of some components in leaves and bulbs of Spanish bluebell (Hyacinthoides hispanica (Mill.) Rothm.). J. Elem., 24(1): 305-318. DOI: 10.5601/jelem.2018.23.1.1646 RECEIVED: 12 March 2018 ACCEPTED: 23 August 2018 ORIGINAL PAPER INFLUENCE OF MULCHING AND FOLIAR NUTRITION ON THE FORMATION OF BULBS AND CONTENT OF SOME COMPONENTS IN LEAVES AND BULBS OF SPANISH BLUEBELL (HYACINTHOIDES HISPANICA (MILL.) ROTHM.)* Marzena Błażewicz-Woźniak1, Dariusz Wach1, Agnieszka Najda2, Sylwia Mucha1 1 Department of Cultivation and Nutrition of Plants 2 Department of Vegetable Crops and Medicinal Plants University of Life Sciences in Lublin, Poland ABSTRACT Hyacinthoides hispanica (Mill.) Rothm. is a typical spring geophyte and has been cultivated for a long time in gardens as an ornamental bulbous plant. Bulbs are not only the storage and dor- mant organs, but also a means for the plant’s vegetative reproduction. In Poland, due to the climate, the wintering and reproduction of this plant can be a problem. The aim of the study was to determine the effect of foliar nutrition using phosphorous fertilizer and soil mulching on the formation of bulbs and content of some components in leaves and bulbs of H. hispanica. To this end, a field experiment was carried out in 2010-2012. Mulching the soil with pine bark had a beneficial effect on the weight of bulbs in the core, the average weight of one bulb and the weight, length, diameter and circumference of the largest bulb from the H. -

Boophone Disticha
Micropropagation and pharmacological evaluation of Boophone disticha Lee Cheesman Submitted in fulfilment of the academic requirements for the degree of Doctor of Philosophy Research Centre for Plant Growth and Development School of Life Sciences University of KwaZulu-Natal, Pietermaritzburg April 2013 COLLEGE OF AGRICULTURE, ENGINEERING AND SCIENCES DECLARATION 1 – PLAGIARISM I, LEE CHEESMAN Student Number: 203502173 declare that: 1. The research contained in this thesis, except where otherwise indicated, is my original research. 2. This thesis has not been submitted for any degree or examination at any other University. 3. This thesis does not contain other persons’ data, pictures, graphs or other information, unless specifically acknowledged as being sourced from other persons. 4. This thesis does not contain other persons’ writing, unless specifically acknowledged as being sourced from other researchers. Where other written sources have been quoted, then: a. Their words have been re-written but the general information attributed to them has been referenced. b. Where their exact words have been used, then their writing has been placed in italics and inside quotation marks, and referenced. 5. This thesis does not contain text, graphics or tables copied and pasted from the internet, unless specifically acknowledged, and the source being detailed in the thesis and in the reference section. Signed at………………………………....on the.....….. day of ……......……….2013 ______________________________ SIGNATURE i STUDENT DECLARATION Micropropagation and pharmacological evaluation of Boophone disticha I, LEE CHEESMAN Student Number: 203502173 declare that: 1. The research reported in this dissertation, except where otherwise indicated is the result of my own endeavours in the Research Centre for Plant Growth and Development, School of Life Sciences, University of KwaZulu-Natal, Pietermaritzburg. -

Regional Landscape Surveillance for New Weed Threats Project 2016-2017
State Herbarium of South Australia Botanic Gardens and State Herbarium Economic & Sustainable Development Group Department of Environment, Water and Natural Resources Milestone Report Regional Landscape Surveillance for New Weed Threats Project 2016-2017 Milestone: Annual report on new plant naturalisations in South Australia Chris J. Brodie, Jürgen Kellermann, Peter J. Lang & Michelle Waycott June 2017 Contents Summary .................................................................................................................................... 3 1. Activities and outcomes for 2016/2017 financial year .......................................................... 3 Funding .................................................................................................................................. 3 Activities ................................................................................................................................ 4 Outcomes and progress of weeds monitoring ........................................................................ 6 2. New naturalised or questionably naturalised records of plants in South Australia. .............. 7 3. Description of newly recognised weeds in South Australia .................................................. 9 4. Updates to weed distributions in South Australia, weed status and name changes ............. 23 References ................................................................................................................................ 28 Appendix 1: Activities of the -
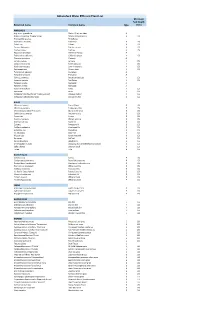
Abbotsford Water Efficient Plant List Minimum Soil Depth Botanical Name Common Name Type (Mm)
Abbotsford Water Efficient Plant List Minimum Soil Depth Botanical name Common name type (mm) ANNUALS Argemone grandifolia Statice/Sea Lavendar A Begonia x hybrida 'Dragon Wings' Dragon Wing begonia A 150 Bracteantha species Strawflower Calendula officinalis Calendula A 150 Coleus ssp. Coleus A 150 Cosmos bipinnatus Garden cosmos A 150 Cuphea llavea Cuphea A 150 Dyssocua tenuiloba Dahlberg Daisey A Eschscholzia californica California poppy A 150 Gazania spendens Gazania A Lantana camara Lantana A 150 Lobularia maritima Sweet alyssum A 150 Nigella damascena Love-in-the-mist A 150 Oesteopermum African daisy A 150 Pelargonium species Geranium A Portulaca oleracea Portulaca A Salvia guaranitica Anise-scented sage A 150 Scaevola aemula Fan flower A 150 Targetes erecta Marogold A Targetes erecta Marogold A Viola x wittrockiana Pansy A 150 Zinnia ssp. Zinnia A 150 Verbascum bombyciferum 'Arctic Summer' Broussa mullein A 150 Verbascum phlomoides 'Spica' Orange mullein A 150 BULBS Allium christophii Star of Persia B 150 Allium karataviense Turkestan onion B 150 Chionodoxa forbesii 'Pink Giant' Glory of the snow B 150 Colchicum autumnale Autumn crocus B 150 Crocus ssp. Crocus B 150 Eranthis hyemalis Winter aconite B 150 Erythronium ssp. Fawn lily B 150 Eucomis Pineapple lily B 150 Fritillaria meleagris Checkered lily B 150 Galanthus ssp. Snowdrop B 150 Iris reticulata Dwarf iris B 150 Muscari ssp. Grape hyacinth B 150 Narcissus Daffodil B 150 Nerine bowdenii Bowden lily B 150 Ornithogalum nutans Drooping Star of Bethlehem/Silverbells B 150 Scilla -
Field Grown Cut Flowers
Nursery FACTSHEET September 2015 Field Grown Cut Flowers INTRODUCTION The culture of field grown flowers is an area of floriculture that is generating a lot of interest and is enjoying a steady growth rate. It provides a way to enter the floriculture industry without the $100 to $150 per square metre capital costs that are involved in some greenhouse crops. Recently, the largest area of growth has been in the specialty cut flowers as opposed to the more traditional field grown crops like statice, dahlias and gypsophila. As gardening increases in popularity, home consumers are becoming familiar with the many new and different flower species. In turn, consumers are starting to look for and demand these flowers in floral design work. Site Selection Whether you plan to lease or own the land, there are basic, yet important, site considerations (see Table 1). It is easier if you start with a suitable site rather than try to modify it later. Table 1. Considerations when selecting a production site Soil: It should be fertile and well drained. Soil tests are a basic management tool. Even if you are familiar with the soil in the area, it must be tested to determine pH, organic matter and nutrient levels. A pH of 6.0–6.5 is suitable for most cuts. Know the requirements of your crop before you make any major changes. Water: Good quality water must be available in sufficient quantities. Have the water source tested to determine essentials like pH and EC (salinity). Terrain: Flat land is easier to work. Watch out for low lying pockets that might be prone to early and late frosts, or flooding during the wet months. -

United (& States • Department of Agricbltm
UNITED STATES DEPARTMENT OF AGRICBLTM • (& INVENTORY No. 75 Washington, D. c. T Issued February, 1926 SEEDS AND PLANTS IMPORTED BY THE OFFICE OF FOREIGN SEED AND PLANT INTRODUCTION, BUREAU OF PLANT INDUSTRY, DURING THE PERIOD FROM APRIL 1 TO JUNE 30,1923 (NOS. 56791 TO 57679) CONTENTS Page Introductory statement 1 Inventory 3 Index of common and scientific names __ 32 INTRODUCTORY STATEMENT When the first Inventory of Seeds and Plants Imported was prepared in 1898, there were practically no government plant-breeding institutions in existence, and almost all of the plants introduced were for direct trial as new crops. Few wild forms were represented, and almost no collections of seeds which were the result of the hybridization or selection work of foreign plant breeders. To-day, as is particularly evident in this inventory, an exchange between the plant breeders of the world is going on which shows a remarkable activity in this field. This practice should be encouraged, for it opens up a wide field of trial for any new variety, and it can be confidently predicted that out of these newly made and plastic forms are likely to come many great commercial varieties of the future. Forms which in the country of their origin have proved inferior to others may prove superior in some other environment. This inventory contains a record of many selected and previously studied varieties of plants sent by foreign plant-breeding institutions: A collection of peanut varieties from the Department of Agriculture at Buitenzorg, Java (Arachis hypogaea; Nos. 56842 to 56849); a new strain of red clover from Dr. -

David Domoneys Complete Guide to Seeds and Bulbs
Do David money’s COMPLETE GUIDE To bs Seeds and Bul Let’s begin at the beginning Knowing where plants come from and how they grow is important if you want to grow strong, healthy plants. Understanding seeds and bulbs will help you become a better gardener. What’s in this guide? First we will look at seeds and the different techniques for sowing them indoors and directly outside. We will also explore how to care for seedlings, plus the right ways to collect and store seeds from your plants. In the second part, we will talk about the different types of bulbs, how to plant them and the best ways to create year-round displays. Seeds Growing your own plants from seed is always exciting. Seeing them develop into new plants is very rewarding, and it’s a cost-effective way to add to your garden. What are seeds? A seed is the unit of reproduction for most plants. It contains all the genetic information needed to create an entirely new plant, plus a nutrient store to help it get started. So many types of plants can be grown from seed, from herbs and vegetables to annual and perennial flowering species. Most seeds need warmth and moisture to germinate, but some need bright light or a cooler environment to get started. Others require soaking before sowing. There will be full instructions on the seed packet, but here I will show you a few general tips to help you get started. Find more guides at davidomoney.com Sowing outdoors Some seeds can be sown directly into the ground where you want them to grow. -

Nerine Bowdenii Final Trials Report 2012–2017
Nerine bowdenii Final Trials Report 2012–2017 Trials Office, RHS Garden Wisley, Woking, Surrey, GU23 6QB RHS Registered Charity No: 222879 / SC038262 RHS Trial of Nerine bowdenii Introduction to the Trial The genus Nerine contains 25 species, most well known of which is the Guernsey lily (N. sarniensis) a greenhouse plant. All the species occur in southern Africa but only a very few are truly hardy in the UK. Since its introduction to cultivation in 1898 N. bowdenii has become widely grown in gardens and a diverse range of forms has been developed, either by selection or hybridisation. The scale of this diversity was revealed by the Hardy Nerine Study Day in October 2007. From that point onwards the idea of a trial grew, mainly through the efforts of the then National Plant Collection holder for hardy nerines, Mrs Margaret Owen. The trial agreed was an RHS off-site trial, run jointly by the RHS and the Nerine and Amaryllid Society (NAAS). Two sites were chosen to test the hardiness of the entries: one in the north (The Patch, Acton Pigott, Shropshire – site of the then National Plant Collection) and one in the south (Bramdean House, Bramdean, Hampshire – site of an extensive collection of nerines owned by Mrs Victoria Wakefield). Although the intention was to have entries in common to both sites to allow proper comparison, the difficult process of putting the trial together meant that not all the entries were grown at both sites. A particular challenge for the trial was the fact that many of the entries were not named or not properly named as cultivars. -

Accumulation of Heavy Metals in the Organs of Herbaceous Plants
E3S Web of Conferences 265, 02019 (2021) https://doi.org/10.1051/e3sconf/202126502019 APEEM 2021 Accumulation of heavy metals in the organs of herbaceous plants Antonina Reut, Aigul Biglova, and Irina Allayarova South-Ural Botanical Garden-Institute of Ufa Federal Research Centre of Russian Academy of Sciences, 450080, Mendeleev str., 195/3, Ufa, Russia Abstract. The purpose of the work is to establish the characteristics of the accumulation of heavy metals of aboveground and underground organs in plant samples of 8 varieties of Narcissus hybridus and Camassia cusickii in the urbanized environment of the city of Ufa. The study of the elemental composition of the samples was carried out according to the method of quantitative chemical analysis ‘Determination of As, Pb, Cd, Sn, Cr, Cu, Fe, Mn and Ni in laboratory samples of food products and food raw materials by the atomic absorption method with electrothermal atomization’. It was found that the content of nickel, copper, cadmium, lead, magnesium, iron, chromium in all the samples studied does not exceed the standards specified in the State Pharmacopoeia of the Russian Federation (SP RF). The arsenic content exceeds the standards specified in the SP RF by 3.4 times. Revealed a high intensity of biological absorption of copper. It has been established that the concentrations of As, Cu, Pl, Mn, Fe in the aboveground organs are higher than in the underground part of plants; the ratio of the content of Ni, Cr, Cd in aboveground and underground organs is the same. 1 Introduction The ecological state of the urban environment has a significant impact on human life. -

Nerine & Amaryllid Society
Copyright of The Nerine and Amaryllid Society Nerine & Amaryllid Society Amaryllids-the Journal of the Nerine & Amaryllid Society. Index to Journals 1998-2017 Re-prints are available to members. Individual Journals can be supplied free as PDF files by e-mail; printed at a charge of £2 each. Available from; Andrew Eames, 01332 792569, [email protected] Caroline Stone, (PDF only) 01566 785706, [email protected] If you want many please contact one of us to discuss. www.nerineandamaryllidsociety.co.uk 16 Vol. 1 No.1 1998 2017 Part 3 Nerine in the New Zealand of the 1970s Roger Beauchamp Hoyland Plant Nursery Visit Jonny Hartnell Cyrtanthus.-neglected by Nerine Enthusiasts? Paul Chapman The smaller Snowflakes Dr John David Society aims and objectives Roger Beauchamp The Godman family & Nerines Malcolm Allison Vol.1 No.2. Developments at Bickham Cottage Steve Eyre Echoes from the past Roger Beauchamp RHS Nerine bowdenii Trial update Mark Heath Scent in the Genus Nerine. HJ Dalton RHS Allium Trial Report Karen Robbirt Why don’t my Nerines flower? The Dutch way- 1957 RHS Hippeastrum Trial Report Karen Robbirt The English way-1966. Boophone disticha Mike Jeans Nerines in the United States Charles Hardman Bountiful Belladonnas Bradley Newton Vol.1 No.3. Wildside Agapanthus Caroline Stone The start of interest in Nerine. Marion Wood Memories of Stanley Smee. Gerald W Goddard Hippeastrum Developments. Veronica Read South African Bulbs. Terry Smale Vol.1 No.4. The fascinating world of Nerines. LL Mead Lycoris Roger Beauchamp on the cultivation of Nerines (Oct 1970) Stanley Smee Vol.2 No.1 1999 The bi-centenary of the Hybrid Hippeastrum. -

Going to Sustainable
GOING TO SUSTAINABLE Lowering Landscape and Garden Maintenance Including Better Ways to Water and How to Save Water © Joseph L. Seals, 2008, 2009 Copyright Joseph L. Seals, 2008, 2009 LOWERING MAINTENANCE REDUCING MAINTENANCE IN THE PLANNING STAGES Unfortunately, maintenance of the landscape is often assumed or overlooked during the planning and design phase of a project 1) Keep the planting design simple. The more elaborate the plan and planting -- Numbers of plants, variety of plants, -- less than simple lines and shapes -- … the more maintenance is required. For instance, lawn areas need to be plotted so that mowing, edging and periodic maintenance can be accomplished easily. -- Avoid tight angles and sharp corners. -- wide angles, gentle, sweeping curves, and straight lines are much easier to mow. -- Make certain each plant in the plan serves a purpose. 2) Select the right plant for the right place We all know that there are “sun plants” for sunny spots and “shade plants” for shady spots. And we don’t plant “sun plants” in shade nor do we plant “shade plants” in sun. And some of us know that there are drought-tolerant plants that like dry soil and little water -- and there are moisture-loving plants that like their feet wet. And we don’t mix those up either. Such “mix ups” result in everything from the obvious: outright death of the plant involved to a subtly stressed plant that shows various symptoms of “disease” -- whether it’s an actual organism or a physiological condition. Copyright Joseph L. Seals, 2008, 2009 Every time you push a plant beyond its natural adaptations, abilities, and tolerances, you invite problems and you invite higher maintenance When choosing the right plant, start with THE BIG PICTURE: We have a Mediterranean climate.