Diet Breadth, Coexistence and Rarity in Bumblebees
Total Page:16
File Type:pdf, Size:1020Kb

Load more
Recommended publications
-

UNIVERSITY of READING Delivering Biodiversity and Pollination Services on Farmland
UNIVERSITY OF READING Delivering biodiversity and pollination services on farmland: a comparison of three wildlife- friendly farming schemes Thesis submitted for the degree of Doctor of Philosophy Centre for Agri-Environmental Research School of Agriculture, Policy and Development Chloe J. Hardman June 2016 Declaration I confirm that this is my own work and the use of all material from other sources has been properly and fully acknowledged. Chloe Hardman i Abstract Gains in food production through agricultural intensification have come at an environmental cost, including reductions in habitat diversity, species diversity and some ecosystem services. Wildlife- friendly farming schemes aim to mitigate the negative impacts of agricultural intensification. In this study, we compared the effectiveness of three schemes using four matched triplets of farms in southern England. The schemes were: i) a baseline of Entry Level Stewardship (ELS: a flexible widespread government scheme, ii) organic agriculture and iii) Conservation Grade (CG: a prescriptive, non-organic, biodiversity-focused scheme). We examined how effective the schemes were in supporting habitat diversity, species diversity, floral resources, pollinators and pollination services. Farms in CG and organic schemes supported higher habitat diversity than farms only in ELS. Plant and butterfly species richness were significantly higher on organic farms and butterfly species richness was marginally higher on CG farms compared to farms in ELS. The species richness of plants, butterflies, solitary bees and birds in winter was significantly correlated with local habitat diversity. Organic farms supported more evenly distributed floral resources and higher nectar densities compared to farms in CG or ELS. Compared to maximum estimates of pollen demand from six bee species, only organic farms supplied sufficient pollen in late summer. -
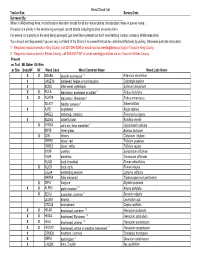
Weed Check List Survey Date
Weed Check List Trail or Site: Survey Date: Surveyed By: When in Wildnerness Area, record location and other details for all non-native plants. Include plant name in survey notes. If a weed is a priority in the area being surveyed, record details including location on survey form. For weeds not a priority in the area being surveyed, just mark them present but don't need lat/long location unless in Wilderness Area. You can pull and bag weeds if you are very confident of the ID and it is a weed that can be controlled effectively by pulling. Otherwise just note its location. 1 - Regulated noxious weeds in King County, call 206-296-0290 or email [email protected] if found in King County 2 - Regulated noxious weed in Kittitas County, call 509-962-7007 or email [email protected] if found in Kittitas County Present on Trail Mt. Baker Ok Wen or Site Snoq NF NF Weed Code Weed Common Name Weed Latin Name XXARAB3 absinth wormwood 1,2 Artemisia absinthium X CASE13 bindweed, hedge or morning glory Calystegia sepium X SODU bittersweet nightshade Solanum dulcamara XXRULA blackberry, evergreen or cutleaf 2 Rubus laciniatus XXRUAR9 blackberry, Himalayan 2 Rubus armeniacus SILA21 bladder campion 2 Silene latifolia X AJRE bugleweed Ajuga reptans RARE3 buttercup, creeping Ranunculus repens X BUDA2 butterfly bush Buddleia davidii X HYRA3 cat’s ear, false dandelion 2 Hypochaeris radicata BRTE cheat grass Bromus tectorum X CIIN chicory Cichorium intybus TRPR2 clover, red Trifolium pratense TRRE3 clover, white Trifolium repens SYOF comfrey Symphytum officinale TAOF dandelion Taraxacum officinale RUOB dock, broadleaf Rumex obtusifolius X RUCR dock, curly Rumex crispus LALA4 everlasting peavine Lathyrus latifolius MAPE2 false mayweed Tripleurospermum perforatum X DIPU foxglove Digitalis purpurea XXALPE4 garlic mustard 1,2 Alliaria petiolata X SEVU groundsel, common 2 Senecio vulgaris LEONT hawkbit Leontodon spp. -

Sterol Addition During Pollen Collection by Bees
Sterol addition during pollen collection by bees: another possible strategy to balance nutrient deficiencies? Maryse Vanderplanck, Pierre-Laurent Zerck, Georges Lognay, Denis Michez To cite this version: Maryse Vanderplanck, Pierre-Laurent Zerck, Georges Lognay, Denis Michez. Sterol addition during pollen collection by bees: another possible strategy to balance nutrient deficiencies?. Apidologie, 2020, 51 (5), pp.826-843. 10.1007/s13592-020-00764-3. hal-02784696 HAL Id: hal-02784696 https://hal.archives-ouvertes.fr/hal-02784696 Submitted on 3 May 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Apidologie (2020) 51:826–843 Original article * INRAE, DIB and Springer-Verlag France SAS, part of Springer Nature, 2020 DOI: 10.1007/s13592-020-00764-3 Sterol addition during pollen collection by bees: another possible strategy to balance nutrient deficiencies? 1,2 1 3 1 Maryse VANDERPLANCK , Pierre-Laurent ZERCK , Georges LOGNAY , Denis MICHEZ 1Laboratory of Zoology, Research Institute for Biosciences, University of Mons, 20 Place du Parc, 7000, Mons, Belgium 2CNRS, UMR 8198 - Evo-Eco-Paleo, Univ. Lille, F-59000, Lille, France 3Analytical Chemistry, Agro Bio Chem Department, Gembloux Agro-Bio Tech University of Liège, 2 Passage des Déportés, 5030, Gembloux, Belgium Received 10 July 2019 – Revised2March2020– Accepted 30 March 2020 Abstract – Sterols are essential nutrients for bees which are thought to obtain them exclusively from pollen. -

Shoot Structure of Symphytum Officinale L. (Boraginaceae) in Relation to the Nature of Its Axially Shifted Lateral Branches
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Wulfenia Jahr/Year: 2011 Band/Volume: 18 Autor(en)/Author(s): Kotelnikova Ksenia V., Tretjakova Daria J., Nuraliev Maxim S. Artikel/Article: Shoot structure of Symphytum officinale L. (Boraginaceae) in relation to the nature of its axially shifted lateral branches. 63-79 © Landesmuseum für Kärnten; download www.landesmuseum.ktn.gv.at/wulfenia; www.biologiezentrum.at Wulfenia 18 (2011): 63 –79 Mitteilungen des Kärntner Botanikzentrums Klagenfurt Shoot structure of Symphytum offi cinale L. (Boraginaceae) in relation to the nature of its axially shifted lateral branches Ksenia V. Kotelnikova, Daria J. Tretjakova & Maxim S. Nuraliev Summary: Shoot structure of Symphytum offi cinale was studied in terms of its architecture, vascular anatomy and morphogenesis. The main stem and lower stems of secondary order typically bear axially shifted lateral branches (along with axillary ones). This results in development of internode-like stem section between the lateral branch and its subtending leaf. Each shifted branch possesses two stem leaves (prophylls) and a terminal infl orescence. All lateral buds are initiated in the leaf axils. At later developmental stages some of them become shifted due to the intercalary growth of the stem between them and their subtending leaves. The intercalary stem portion possesses internodal stele structure, and the stele of the lateral stem branches from the main stele almost at the level of lateral stem insertion. This phenomenon can be treated as concaulescence (i.e. congenital fusion), though no strict evidences for phylogenetic fusion were found. Stems of Symphytum offi cinale also bear green wings of leafy nature. -

Boraginaceae), and the Phylogeny of Boraginoideae
!" #$ % " "& '()*"'+ (,-./01 ** -)2'/)*) %*()'-) %%*(* 3443 Dissertation for the Degree of Doctor of Philosophy in Systematic Botany presented at Uppsala University in 2002 Abstract Långström, E. 2002. Systematics of Echiochilon and Ogastemma (Boraginaceae), and the phylogeny of Boraginoideae. Acta Univ. Ups. Comprehensive Summaries og Uppsala Disserta- tions from the Faculty of Science and Technology 693. 34 pp. Uppsala. ISBN 91-554-5257-4. Echiochilon, Ogastemma and Sericostoma are revised resulting in the recognition of 15 spe- cies of Echiochilon and one Ogastemma species. Several species are placed in synonymy and three new species are described, E. baricum, E. callianthum and E. cyananthum. The single species of Sericostoma is shown to be nested within Echiochilon. The plastid atpB gene was sequenced for Echiochilon and Ogastemma from the Old World and Antiphytum from the New World, plus for a selection of 33 other Boraginaceae taxa. They were analysed together with selected outgroup taxa to give a framework of the tribes of Boraginoi- deae. The analysis gave support for establishing the new tribe Echiochileae for Antiphytum, Echio- chilon and Ogastemma, and for merging the traditionally accepted tribe Eritrichieae with Cyno- glosseae. The ITS region was sequenced for all but one species of Echiochilon and for representa- tives of Antiphytum and Ogastemma. Phylogenetic analysis of Echiochilon revealed that the strongly zygomorphic-flowered species form a paraphyletic group. The morphological data gave results fairly congruent with the ITS phylogeny. Biogeographic interpretations of the ITS and atpB phylogenies indicated a trans-Atlantic dispersal of Antiphytum as the most plausible explanation to the Old/New World disjunction. Analyses using DIVA (Dispersal Vicariance Analysis) of the distributions of the Echiochilon spe- cies indicated an ancestor to Echiochilon with a wide distribution over northern Africa and Arabia to India. -

Guidebook to Invasive Nonnative Plants of the Elwha Watershed Restoration
Guidebook to Invasive Nonnative Plants of the Elwha Watershed Restoration Olympic National Park, Washington Cynthia Lee Riskin A project submitted in partial fulfillment of the requirements for the degree of Master of Environmental Horticulture University of Washington 2013 Committee: Linda Chalker-Scott Kern Ewing Sarah Reichard Joshua Chenoweth Program Authorized to Offer Degree: School of Environmental and Forest Sciences Guidebook to Invasive Nonnative Plants of the Elwha Watershed Restoration Olympic National Park, Washington Cynthia Lee Riskin Master of Environmental Horticulture candidate School of Environmental and Forest Sciences University of Washington, Seattle September 3, 2013 Contents Figures ................................................................................................................................................................. ii Tables ................................................................................................................................................................. vi Acknowledgements ....................................................................................................................................... vii Introduction ....................................................................................................................................................... 1 Bromus tectorum L. (BROTEC) ..................................................................................................................... 19 Cirsium arvense (L.) Scop. (CIRARV) -

Brittlestem Hempnettle Galeopsis Tetrahit L
splitlip hempnettle Galeopsis bifida Boenn. brittlestem hempnettle Galeopsis tetrahit L. Synonyms for Galeopsis bifida: Galeopsis bifida var. emarginata Nakai, G. tetrahit var. bifida (Boenn.) Lej. & Court., G. tetrahit var. parviflora Bentham Other common name: none Synonyms for Galeopsis tetrahit: none Other common name: common hempnettle Family: Lamiaceae Invasiveness Rank: 50 The invasiveness rank is calculated based on a species’ ecological impacts, biological attributes, distribution, and response to control measures. The ranks are scaled from 0 to 100, with 0 representing a plant that poses no threat to native ecosystems and 100 representing a plant that poses a major threat to native ecosystems. Description These Galeopsis species can be distinguished from each Splitlip hempnettle and brittlestem hempnettle are other based on the following features. Brittlestem annual plants that grow 20 to 80 cm tall from taproots. hempnettle has stems that are hairy mainly on the nodes, Stems are erect, quadrangular, simple or branched, rounded leaf bases, and central lobes on the corollas that hairy, and usually swollen below nodes. Leaves are are entire. Corollas of brittlestem hempnettle are 15 to opposite, petiolated, lanceolate to ovate, sparsely hairy 23 mm long. Splitlip hempnettle has internodes that are on both sides, 3 to 10 cm long, and 1 to 5 cm wide with usually densely hairy, triangular leaf bases, and central long-pointed tips and coarsely toothed margins. Petioles lobes on the corollas that are notched. Corollas of are hairy and 1 to 2.5 cm long. Flowers are arranged in splitlip hempnettle are usually less than 15 mm long dense clusters in leaf axils near the ends of stems. -

New Jersey Strategic Management Plan for Invasive Species
New Jersey Strategic Management Plan for Invasive Species The Recommendations of the New Jersey Invasive Species Council to Governor Jon S. Corzine Pursuant to New Jersey Executive Order #97 Vision Statement: “To reduce the impacts of invasive species on New Jersey’s biodiversity, natural resources, agricultural resources and human health through prevention, control and restoration, and to prevent new invasive species from becoming established.” Prepared by Michael Van Clef, Ph.D. Ecological Solutions LLC 9 Warren Lane Great Meadows, New Jersey 07838 908-637-8003 908-528-6674 [email protected] The first draft of this plan was produced by the author, under contract with the New Jersey Invasive Species Council, in February 2007. Two subsequent drafts were prepared by the author based on direction provided by the Council. The final plan was approved by the Council in August 2009 following revisions by staff of the Department of Environmental Protection. Cover Photos: Top row left: Gypsy Moth (Lymantria dispar); Photo by NJ Department of Agriculture Top row center: Multiflora Rose (Rosa multiflora); Photo by Leslie J. Mehrhoff, University of Connecticut, Bugwood.org Top row right: Japanese Honeysuckle (Lonicera japonica); Photo by Troy Evans, Eastern Kentucky University, Bugwood.org Middle row left: Mile-a-Minute (Polygonum perfoliatum); Photo by Jil M. Swearingen, USDI, National Park Service, Bugwood.org Middle row center: Canadian Thistle (Cirsium arvense); Photo by Steve Dewey, Utah State University, Bugwood.org Middle row right: Asian -

Phylogeny of Eudicots (Or Tricolpates) Eudicots (Or Tricolpates) “Basal Eudicots”
Phylogeny of Eudicots (or Tricolpates) Eudicots (or Tricolpates) “Basal eudicots” Asterids Buxales Rosids Caryophyllales RanunculalesProteales After Jansen et al., 2007, Proc. Natl. Acad. Sci. USA 104: 19369-19374 Phylogeny of Asterids Asterids Lamiids Ericales Campanulids Cornales Cornales Ericales Campanulids: Lamiids: Aquifoliales Garryales Apiales Gentianales Dipsacales Lamiales Asterales Solanales After APG, 2003; Judd and Olmstead, 2004, and Soltis et al., 2005 Synapomorphies for Asterids and Core Asterids Synapomorphies for Asterids: Iridoid compounds; Unitegmic ovules; www.scielo.br/img/fbpe/jbchs/v12n2/a04fig01.gif Tenuinucellate ovules. Synapomorphies for Core Asterids (Lamiids + Campanulids): Gamopetalous corollas; A single whorl of stamens that alternate with the petal lobes; Epipetalous stamens; 2 fused carpels. Ericaceae (Rhododendron family) http://georgian.wunderground.com/data/wximagenew/f/Feather3/5431.jpg Rhododendron macrophyllum (Pacific Rhododendron, WA State Flower) Ericaceae (Rhododendron family) Textbook DVD WSJ Textbook DVD KRR & DLN Vaccinium corymbosum; The genus Vaccinium contains about 450 species, including blueberry, cranberry, huckleberry, etc.. Ericaceae (Rhododendron family) Photo: Yaowu Yuan Photo: Yaowu Yuan Gautheria shallon; Salal Cassiope tetragona http://www.nestlerode.org/ Cremation_Urn/urn_finished.jpg Photo: Yaowu Yuan Note the more or less pendulous Menziesia ferruginea flowers and the urn-shaped corolla. Ericaceae (Rhododendron family) Textbook DVD KRR & DLN Textbook DVD KMN Pieris japonica Vaccinium arboreum Note that the anthers often have spur- or awn-like appendages. Ericaceae (Rhododendron family) Photo: Yaowu Yuan Menziesia ferruginea Note that anther opens by 2 terminal pores –– poricidal dehiscence. Ericaceae (Rhododendron family) Textbook DVD KRR & DLN Kalmia latifolia Note that although corolla can be more spread and not urn-shaped, anther usually opens by terminal pores. -

Utilisation of Pollen Resources by Bumblebees in an Enhanced Arable Landscape
Final contract report to English Nature, Project VT0312 22nd February 2004 Utilisation of pollen resources by bumblebees in an enhanced arable landscape Claire Carvell, Bill Meek, Richard Broughton, Tim Sparks and Richard Pywell NERC Centre for Ecology and Hydrology, Monks Wood, Abbots Ripton, Huntingdon, PE28 2LS, UK. Tel: 01487 772400 Paul Westrich Lichtensteinstr. 17, D-72127 Kusterdingen, Germany Marek Nowakowski The Farmed Environment Company, Manor Farm, Eddlethorpe, York, North Yorkshire YO17 9QT, UK Summary · Pollen is an essential resource for bumblebees, yet this is often overlooked in studies of their foraging requirements. The decline of bumblebees across the UK and rest of Europe has prompted conservation measures to consider the provision of pollen as well as nectar resources, particularly in intensive agricultural landscapes where the pollination service bumblebees provide is so important. · This report details a study carried out in July 2003 to investigate the utilisation of pollen from different forage plant species by two bumblebee species with contrasting ecologies (Bombus pascuorum and Bombus terrestris/lucorum) across an enhanced arable landscape. · An area of 1.96 km2, centred on Manor Farm, near Malton in North Yorkshire, was divided into grid squares of 100m x 100m. Pollen loads were sampled from foraging bumblebees in eight random squares within the centre of this grid, and every square was surveyed in detail to map the distribution and abundance of all plant species in flower. Each pollen load was analysed to identify the pollen genera and/or species present, and to estimate the percentage species composition of the load. · The two bumblebee species showed specialization towards pollen from contrasting species. -

Boraginaceae) Taxa from Turkey
Bangladesh J. Bot. 36(2): 93-103, 2007 (December) MICROMORPHOLOGY AND ANATOMY OF THREE SYMPHYTUM (BORAGINACEAE) TAXA FROM TURKEY OZNUR ERGEN AKCIN AND HILAL BAKI Department of Biology, Sciences & Arts Faculty, Ordu University, Ordu, Turkey E-mail: [email protected] Key words: Micromorphology, Anatomy, Symphytum Abstract Symphytum asperum Lepechin, S. ibericum Steven and S. sylvaticum Boiss. were examined morphologically, micromorphologically and anatomically. Scanning electron microscopy was used to examine leaf surface and trichomes of these species. These species had bifacial and hypostomatous leaf types. Epidermal cells of leaves were usually polygonal or irregular in form. The pattern of anticlinical cells may vary in different species and between the upper and lower epidermis of the same species. Stomata are anisocytic and anomocytic in three species. Stomata index is 27.5 for S. sylvaticum, 24.65 for S. ibericum and 21.86 for S. asperum glandular trichomes are capitate in forms and more dense on the lower epidermis than upper epidermis. Eglandular trichomes are simple, short or long, unicellular or multicellular and thin or thick. Introduction Symphytum L. (Boraginaceae) is represented by about 23 taxa in Turkey (Wickens 1978, Yildirimli 2000). Symphytum species are common garden plants and have been used as a source of herbal medicines for >2000 years. Leaf and root of Symphytum species are used by lay public, herbalists and physicians for treatment of broken bones, tendon damage, ulcerations in the gastrointestinal tract, and lung congestion. These species are also rich in many crucial nutrients, such as protein, antioxidant, vitamins and vitamin B12, and are common component in the diet of certain ethnic groups (Hills 1976, Rode 2002). -

Invasive Plant Inventory and Bird Cherry Control Trials
MUNICIPALITY OF ANCHORAGE NON-NATIVE PLANT SURVEY A survey of non-native plants along major and secondary roads in the Municipality of Anchorage, Alaska February 29, 2012 MUNICIPALITY OF ANCHORAGE NON-NATIVE PLANT SURVEY A survey of non-native plants along major and secondary roads in the Municipality of Anchorage, Alaska Prepared for: Anchorage Park Foundation and Municipality of Anchorage Anchorage Park Foundation 715 L Street, Suite 200 Anchorage, AK 99501 Prepared by: Helen Klein, Casey Greenstein, Miriah Phelps, Lindsey Flagstad, Helen Cortés-Burns and Matthew Carlson Alaska Natural Heritage Program University of Alaska Anchorage 707 A Street Anchorage, AK 99501 February 29, 2012 MUNICIPALITY OF ANCHORAGE NON-NATIVE PLANT SURVEY i Table of Contents LIST OF FIGURES .................................................................................................................................................. IV LIST OF TABLES .................................................................................................................................................... VI ABSTRACT ............................................................................................................................................................. 1 ACKNOWLEDGEMENTS ........................................................................................................................................... 2 INTRODUCTION ......................................................................................................................................................