Boraginaceae) Taxa from Turkey
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

State of New York City's Plants 2018
STATE OF NEW YORK CITY’S PLANTS 2018 Daniel Atha & Brian Boom © 2018 The New York Botanical Garden All rights reserved ISBN 978-0-89327-955-4 Center for Conservation Strategy The New York Botanical Garden 2900 Southern Boulevard Bronx, NY 10458 All photos NYBG staff Citation: Atha, D. and B. Boom. 2018. State of New York City’s Plants 2018. Center for Conservation Strategy. The New York Botanical Garden, Bronx, NY. 132 pp. STATE OF NEW YORK CITY’S PLANTS 2018 4 EXECUTIVE SUMMARY 6 INTRODUCTION 10 DOCUMENTING THE CITY’S PLANTS 10 The Flora of New York City 11 Rare Species 14 Focus on Specific Area 16 Botanical Spectacle: Summer Snow 18 CITIZEN SCIENCE 20 THREATS TO THE CITY’S PLANTS 24 NEW YORK STATE PROHIBITED AND REGULATED INVASIVE SPECIES FOUND IN NEW YORK CITY 26 LOOKING AHEAD 27 CONTRIBUTORS AND ACKNOWLEGMENTS 30 LITERATURE CITED 31 APPENDIX Checklist of the Spontaneous Vascular Plants of New York City 32 Ferns and Fern Allies 35 Gymnosperms 36 Nymphaeales and Magnoliids 37 Monocots 67 Dicots 3 EXECUTIVE SUMMARY This report, State of New York City’s Plants 2018, is the first rankings of rare, threatened, endangered, and extinct species of what is envisioned by the Center for Conservation Strategy known from New York City, and based on this compilation of The New York Botanical Garden as annual updates thirteen percent of the City’s flora is imperiled or extinct in New summarizing the status of the spontaneous plant species of the York City. five boroughs of New York City. This year’s report deals with the City’s vascular plants (ferns and fern allies, gymnosperms, We have begun the process of assessing conservation status and flowering plants), but in the future it is planned to phase in at the local level for all species. -

FDA OTC Reviews Summary of Back Issues
Number 23 The Journal of the AMERICAN BOTANI CAL COUNCIL and the HERB RESEARCH FOUNDATION Chinese Medicinals -A Comprehensive Review of Chinese Materia Medica Legal and Regulatory- FDA OTC Reviews Summary of Back Issues Ongoing Market Report, Research Reviews (glimpses of studies published in over a dozen scientific and technical journals), Access, Book Reviews, Calendar, Legal and Regulatory, Herb Blurbs and Potpourri columns. #1 -Summer 83 (4 pp.) Eucalyptus Repels Reas, Stones Koalas; FDA OTC tiveness; Fungal Studies; More Polysaccharides; Recent Research on Ginseng; Heart Panel Reviews Menstrual & Aphrodisiac Herbs; Tabasco Toxicity?; Garlic Odor Peppers; Yew Continues to Amaze; Licorice O.D. Prevention; Ginseng in Perspec Repels Deer; and more. tive; Poisonous Plants Update; Medicinal Plant Conservation Project; 1989 Oberly #2- Fall/Winter 83-84 (8 pp.) Appeals Court Overrules FDA on Food Safety; Award Nominations; Trends in Self-Care Conference; License Plates to Fund Native FDA Magazine Pans Herbs; Beware of Bay Leaves; Tiny Tree: Cancer Cure?; Plant Manual; and more. Comfrey Tea Recall; plus. #17-Summer 88. (24 pp.) Sarsaparilla, A Literature Review by Christopher #3-Spring 84 (8 pp.) Celestial Sells to Kraft; Rowers and Dinosaurs Demise?; Hobbs; Hops May Help Metabolize Toxins; Herbal Roach Killer; Epazote Getting Citrus Peels for Kitty Litter; Saffron; Antibacterial Sassafras; WHO Studies Anti· More Popular, Aloe Market Levels Off; Herbal Tick Repellent?; Chinese Herb fertility Plants; Chinese Herbal Drugs; Feverfew Migraines; -

Diplomová Práce
Jihočeská univerzita v Českých Budějovicích Zemědělská fakulta Studijní program: N4101 Zemědělské inženýrství Studijní obor: Agroekologie Katedra: Rostlinné výroby a agroekologie Vedoucí katedry: prof. Ing. Vladislav Čurn, Ph.D. DIPLOMOVÁ PRÁCE Biologie, semenářské vlastnosti a zakládání porostů kostivalu lékařského (Symphytum officinale L.) Vedoucí diplomové práce: Ing. Milan Kobes, Ph.D. Autor diplomové práce: Bc. Jaroslava Kovářová České Budějovice, duben 2016 Čestné prohlášení Prohlašuji, že svou diplomovou práci s názvem „Biologie, semenářské vlastnosti a zakládání porostů kostivalu lékařského (Symphytum officinale L.)“ jsem vypracovala samostatně pod vedením vedoucí diplomové práce Ing. Milana Kobese, Ph.D a s použitím odborné literatury a dalších informačních zdrojů, které jsou v práci citovány a zároveň uvedeny v seznamu literatury na konci této práce. Jako autorka uvedené diplomové práce dále prohlašuji, že jsem v souvislosti s jejím vytvořením neporušila autorská práva třetích osob. Dále prohlašuji, že v souladu s § 47b zákona č. 111/1998 Sb. v platném znění souhlasím se zveřejněním své diplomové práce, a to v nezkrácené podobě (v úpravě vzniklé vypuštěním vyznačených částí archivovaných Zemědělskou fakultou JU) elektronickou cestou ve veřejně přístupné části databáze STAG provozované Jihočeskou univerzitou v Českých Budějovicích na jejích internetových stránkách. V Českých Budějovicích dne.......... _____________________________ Poděkování Velké poděkování patří mému vedoucímu práce Ing. Milanu Kobesovi, Ph.D. za odborné rady, připomínky, za vedení a odborné konzultace, které mi poskytl během zpracování této diplomové práce. Děkuji celé své rodině za podporu. ABSTRAKT Diplomová práce s názvem ,,Biologie, semenářské vlastnosti a zakládání porostů kostivalu lékařského (Symphytum officinale L.)“ se zabývala hodnocením způsobů rozmnožování kostivalu a hodnocením vlivu ekologických podmínek na růst kostivalu. Dále testy klíčivosti a jejich statistickým hodnocením při respektování specifik práce se semeny. -
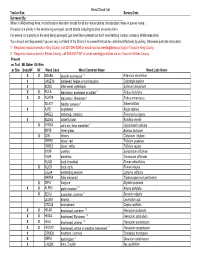
Weed Check List Survey Date
Weed Check List Trail or Site: Survey Date: Surveyed By: When in Wildnerness Area, record location and other details for all non-native plants. Include plant name in survey notes. If a weed is a priority in the area being surveyed, record details including location on survey form. For weeds not a priority in the area being surveyed, just mark them present but don't need lat/long location unless in Wilderness Area. You can pull and bag weeds if you are very confident of the ID and it is a weed that can be controlled effectively by pulling. Otherwise just note its location. 1 - Regulated noxious weeds in King County, call 206-296-0290 or email [email protected] if found in King County 2 - Regulated noxious weed in Kittitas County, call 509-962-7007 or email [email protected] if found in Kittitas County Present on Trail Mt. Baker Ok Wen or Site Snoq NF NF Weed Code Weed Common Name Weed Latin Name XXARAB3 absinth wormwood 1,2 Artemisia absinthium X CASE13 bindweed, hedge or morning glory Calystegia sepium X SODU bittersweet nightshade Solanum dulcamara XXRULA blackberry, evergreen or cutleaf 2 Rubus laciniatus XXRUAR9 blackberry, Himalayan 2 Rubus armeniacus SILA21 bladder campion 2 Silene latifolia X AJRE bugleweed Ajuga reptans RARE3 buttercup, creeping Ranunculus repens X BUDA2 butterfly bush Buddleia davidii X HYRA3 cat’s ear, false dandelion 2 Hypochaeris radicata BRTE cheat grass Bromus tectorum X CIIN chicory Cichorium intybus TRPR2 clover, red Trifolium pratense TRRE3 clover, white Trifolium repens SYOF comfrey Symphytum officinale TAOF dandelion Taraxacum officinale RUOB dock, broadleaf Rumex obtusifolius X RUCR dock, curly Rumex crispus LALA4 everlasting peavine Lathyrus latifolius MAPE2 false mayweed Tripleurospermum perforatum X DIPU foxglove Digitalis purpurea XXALPE4 garlic mustard 1,2 Alliaria petiolata X SEVU groundsel, common 2 Senecio vulgaris LEONT hawkbit Leontodon spp. -

Sterol Addition During Pollen Collection by Bees
Sterol addition during pollen collection by bees: another possible strategy to balance nutrient deficiencies? Maryse Vanderplanck, Pierre-Laurent Zerck, Georges Lognay, Denis Michez To cite this version: Maryse Vanderplanck, Pierre-Laurent Zerck, Georges Lognay, Denis Michez. Sterol addition during pollen collection by bees: another possible strategy to balance nutrient deficiencies?. Apidologie, 2020, 51 (5), pp.826-843. 10.1007/s13592-020-00764-3. hal-02784696 HAL Id: hal-02784696 https://hal.archives-ouvertes.fr/hal-02784696 Submitted on 3 May 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Apidologie (2020) 51:826–843 Original article * INRAE, DIB and Springer-Verlag France SAS, part of Springer Nature, 2020 DOI: 10.1007/s13592-020-00764-3 Sterol addition during pollen collection by bees: another possible strategy to balance nutrient deficiencies? 1,2 1 3 1 Maryse VANDERPLANCK , Pierre-Laurent ZERCK , Georges LOGNAY , Denis MICHEZ 1Laboratory of Zoology, Research Institute for Biosciences, University of Mons, 20 Place du Parc, 7000, Mons, Belgium 2CNRS, UMR 8198 - Evo-Eco-Paleo, Univ. Lille, F-59000, Lille, France 3Analytical Chemistry, Agro Bio Chem Department, Gembloux Agro-Bio Tech University of Liège, 2 Passage des Déportés, 5030, Gembloux, Belgium Received 10 July 2019 – Revised2March2020– Accepted 30 March 2020 Abstract – Sterols are essential nutrients for bees which are thought to obtain them exclusively from pollen. -

Draft Assessment Report on Symphytum Officinale L., Radix
12 July 2011 EMA/HMPC/572844/2009 Committee on Herbal Medicinal Products (HMPC) Assessment report on Symphytum officinale L., radix Based on Article 16d(1), Article 16f and Article 16h of Directive 2001/83/EC as amended (traditional use) Draft Herbal substance(s) (binomial scientific name of the plant, including plant part) Symphytum officinale L., radix Herbal preparation(s) Liquid extract (DER 2:1), extraction solvent 65% (V/V) ethanol Pharmaceutical forms Herbal preparations in semi-solid dosage forms for cutaneous use Rapporteur Assessor(s) Note: This Assessment Report is published to support the release for public consultation of the draft Community herbal monograph on Symphytum officinale L., radix. It should be noted that this document is a working document, not yet fully edited, and which shall be further developed after the release for consultation of the monograph. Interested parties are welcome to submit comments to the HMPC secretariat, which the Rapporteur and the MLWP will take into consideration but no ‘overview of comments received during the public consultation’ will be prepared in relation to the comments that will be received on this assessment report. The publication of this draft assessment report has been agreed to facilitate the understanding by Interested Parties of the assessment that has been carried out so far and led to the preparation of the draft monograph. 7 Westferry Circus ● Canary Wharf ● London E14 4HB ● United Kingdom Telephone +44 (0)20 7418 8400 Facsimile +44 (0)20 7523 7051 E-mail [email protected] Website www.ema.europa.eu An agency of the European Union © European Medicines Agency, 2011. -

Public Statement on the Use of Herbal Medicinal Products Containing Pyrrolizidine Alkaloids (Pas) EMA/HMPC/893108/2011 Page 2/22 30 1
1 06 November 2013 2 EMA/HMPC/893108/2011 3 Committee on Herbal Medicinal Products (HMPC) 4 Public statement on the use of herbal medicinal products 5 containing toxic, unsaturated pyrrolizidine alkaloids (PAs) 6 7 2nd Draft 8 Draft discussed by Working Party on Community monographs and November 2011 Community list (MLWP) January 2012 March 2012 May 2012 Adopted by Committee on Herbal Medicinal Products (HMPC) for release 24 September 2012 for consultation Start of public consultation 25 October 2012 End of consultation (deadline for comments) 15 February 2013 2nd draft agreed by Working Party on Community monographs and July 2013 Community list (MLWP) Coordination with Safety Working Party July - October 2013 2nd draft adopted by Committee on Herbal Medicinal Products (HMPC) 17 September 2013 Start of public consultation 14 November 2013 End of consultation (deadline for comments) 15 February 2014 9 Comments should be provided using this template. The completed comments form should be sent to [email protected] 10 Keywords Herbal medicinal products; HMPC; pyrrolizidine alkaloids 11 7 Westferry Circus ● Canary Wharf ● London E14 4HB ● United Kingdom Telephone +44 (0)20 7418 8400 Facsimile +44 (0)20 7523 7051 E-mail [email protected] Website www.ema.europa.eu An agency of the European Union © European Medicines Agency, 2013. Reproduction is authorised provided the source is acknowledged. 12 Public statement on the use of herbal medicinal products 13 containing toxic, unsaturated pyrrolizidine alkaloids (PAs) 14 15 Table of contents 16 1. Introduction (Problem statement) .......................................................... 3 17 1.1. Pyrrolizidine alkaloids (PAs) ...................................................................................3 18 1.2. -

Comfrey (Symphytum Spp.) As an Alternative field Crop Contributing to Closed Agricultural Cycles in Chicken Feeding
Science of the Total Environment 742 (2020) 140490 Contents lists available at ScienceDirect Science of the Total Environment journal homepage: www.elsevier.com/locate/scitotenv Comfrey (Symphytum spp.) as an alternative field crop contributing to closed agricultural cycles in chicken feeding Michael Oster a, Henry Reyer a, Jonas Keiler b, Elizabeth Ball c, Christina Mulvenna c, Eduard Muráni a, Siriluck Ponsuksili a, Klaus Wimmers a,d,⁎ a Leibniz Institute for Farm Animal Biology (FBN), Wilhelm-Stahl-Allee 2, 18196 Dummerstorf, Germany b Institute of Anatomy, Rostock University Medical Center, Gertrudenstrasse 9, 18057 Rostock, Germany c Agri-Food and Biosciences Institute, Large Park, Hillsborough, Co Down BT26 6DR, UK d Faculty of Agricultural and Environmental Sciences, University Rostock, 18059 Rostock, Germany HIGHLIGHTS GRAPHICAL ABSTRACT • Comfrey as a regionally adapted feed crop could be used in poultry farming. • Intake of comfrey leaves showed no ab- erration of health and tissue integrity. • Comfrey leaves in poultry feed can ef- fectively replace rock mineral supple- ments. • Close nutrient cycles by reactivating phosphorus from over-fertilized soils article info abstract Article history: Local cultivars of comfrey (Symphytum spp.) have been used to cover protein and mineral requirements of farm Received 22 April 2020 animals in low-input systems. Due to its known health-promoting (e.g. allantoin), but also anti-nutritive ingre- Received in revised form 21 June 2020 dients (e.g. pyrrolizidine alkaloids), multidisciplinary approaches are essential in order to quantify the nutritional Accepted 22 June 2020 value and the potential of its use in poultry and farm animals in terms of meeting animal needs, using local re- Available online 27 June 2020 sources as well as remediating over-fertilized soils. -

Vascular Plants of Humboldt Bay's Dunes and Wetlands Published by U.S
Vascular Plants of Humboldt Bay's Dunes and Wetlands Published by U.S. Fish and Wildlife Service G. Leppig and A. Pickart and California Department of Fish Game Release 4.0 June 2014* www.fws.gov/refuge/humboldt_bay/ Habitat- Habitat - Occurs on Species Status Occurs within Synonyms Common name specific broad Lanphere- Jepson Manual (2012) (see codes at end) refuge (see codes at end) (see codes at end) Ma-le'l Units UD PW EW Adoxaceae Sambucus racemosa L. red elderberry RF, CDF, FS X X N X X Aizoaceae Carpobrotus chilensis (Molina) sea fig DM X E X X N.E. Br. Carpobrotus edulis ( L.) N.E. Br. Iceplant DM X E, I X Alismataceae lanceleaf water Alisma lanceolatum With. FM X E plantain northern water Alisma triviale Pursh FM X N plantain Alliaceae three-cornered Allium triquetrum L. FS, FM, DM X X E leek Allium unifolium Kellogg one-leaf onion CDF X N X X Amaryllidaceae Amaryllis belladonna L. belladonna lily DS, AW X X E Narcissus pseudonarcissus L. daffodil AW, DS, SW X X E X Anacardiaceae Toxicodendron diversilobum Torrey poison oak CDF, RF X X N X X & A. Gray (E. Greene) Apiaceae Angelica lucida L. seacoast angelica BM X X N, C X X Anthriscus caucalis M. Bieb bur chevril DM X E Cicuta douglasii (DC.) J. Coulter & western water FM X N Rose hemlock Conium maculatum L. poison hemlock RF, AW X I X Daucus carota L. Queen Anne's lace AW, DM X X I X American wild Daucus pusillus Michaux DM, SW X X N X X carrot Foeniculum vulgare Miller sweet fennel AW, FM, SW X X I X Glehnia littoralis (A. -

Shoot Structure of Symphytum Officinale L. (Boraginaceae) in Relation to the Nature of Its Axially Shifted Lateral Branches
ZOBODAT - www.zobodat.at Zoologisch-Botanische Datenbank/Zoological-Botanical Database Digitale Literatur/Digital Literature Zeitschrift/Journal: Wulfenia Jahr/Year: 2011 Band/Volume: 18 Autor(en)/Author(s): Kotelnikova Ksenia V., Tretjakova Daria J., Nuraliev Maxim S. Artikel/Article: Shoot structure of Symphytum officinale L. (Boraginaceae) in relation to the nature of its axially shifted lateral branches. 63-79 © Landesmuseum für Kärnten; download www.landesmuseum.ktn.gv.at/wulfenia; www.biologiezentrum.at Wulfenia 18 (2011): 63 –79 Mitteilungen des Kärntner Botanikzentrums Klagenfurt Shoot structure of Symphytum offi cinale L. (Boraginaceae) in relation to the nature of its axially shifted lateral branches Ksenia V. Kotelnikova, Daria J. Tretjakova & Maxim S. Nuraliev Summary: Shoot structure of Symphytum offi cinale was studied in terms of its architecture, vascular anatomy and morphogenesis. The main stem and lower stems of secondary order typically bear axially shifted lateral branches (along with axillary ones). This results in development of internode-like stem section between the lateral branch and its subtending leaf. Each shifted branch possesses two stem leaves (prophylls) and a terminal infl orescence. All lateral buds are initiated in the leaf axils. At later developmental stages some of them become shifted due to the intercalary growth of the stem between them and their subtending leaves. The intercalary stem portion possesses internodal stele structure, and the stele of the lateral stem branches from the main stele almost at the level of lateral stem insertion. This phenomenon can be treated as concaulescence (i.e. congenital fusion), though no strict evidences for phylogenetic fusion were found. Stems of Symphytum offi cinale also bear green wings of leafy nature. -

Symphytum Tuberosum Complex in Central Europe: Cytogeography, Morphology, Ecology and Taxonomy
Preslia 88: 77–112, 2016 77 Symphytum tuberosum complex in central Europe: cytogeography, morphology, ecology and taxonomy Symphytum tuberosum ve střední Evropě – cytogeografie, morfologie, ekologie a taxonomie LucieKobrlová1,MichalHroneš1,PetrKoutecký2, Milan Š t e c h2 & Bohumil T r á v n í č e k1 1Department of Botany, Faculty of Science, Palacký University, Šlechtitelů 27, CZ-783 71 Olomouc, Czech Republic, e-mail: [email protected], [email protected], [email protected]; 2Department of Botany, Faculty of Science, University of South Bohemia, Branišovská 1760, CZ-370 05 České Budějovice, Czech Republic, email: [email protected], [email protected] Kobrlová L., Hroneš M., Koutecký P., Štech M. & Trávníček B. (2016): Symphytum tuberosum complex in central Europe: cytogeography, morphology, ecology and taxonomy. – Preslia 88: 77–112. The Symphytum tuberosum complex is a highly polyploid and taxonomically intriguing group. At least eight ploidy levels were recorded previously within this complex. Based on flow cytometric screening of 271 central-European populations, two dominant ploidy levels were revealed: tetraploid (2n = 4x = 32) and widespread dodecaploid (2n = 12x = 96). The tetraploid cytotype is mainly distributed along the southern and south-western margins of the West Carpathians where they abut the Pannonian basin, and found only in Slovakia, the Czech Republic (south-eastern Moravia) and Hungary; our findings represent the first records of this ploidy level for the latter two countries. In contrast, the dodecaploid cytotype occurs throughout the whole area studied. In addition to their geographic distributions, differences between the cytotypes in morphology and habitat requirements were detected using a multivariate morphometric analysis and analysis of a phytosociological database, respectively. -

Boraginaceae), and the Phylogeny of Boraginoideae
!" #$ % " "& '()*"'+ (,-./01 ** -)2'/)*) %*()'-) %%*(* 3443 Dissertation for the Degree of Doctor of Philosophy in Systematic Botany presented at Uppsala University in 2002 Abstract Långström, E. 2002. Systematics of Echiochilon and Ogastemma (Boraginaceae), and the phylogeny of Boraginoideae. Acta Univ. Ups. Comprehensive Summaries og Uppsala Disserta- tions from the Faculty of Science and Technology 693. 34 pp. Uppsala. ISBN 91-554-5257-4. Echiochilon, Ogastemma and Sericostoma are revised resulting in the recognition of 15 spe- cies of Echiochilon and one Ogastemma species. Several species are placed in synonymy and three new species are described, E. baricum, E. callianthum and E. cyananthum. The single species of Sericostoma is shown to be nested within Echiochilon. The plastid atpB gene was sequenced for Echiochilon and Ogastemma from the Old World and Antiphytum from the New World, plus for a selection of 33 other Boraginaceae taxa. They were analysed together with selected outgroup taxa to give a framework of the tribes of Boraginoi- deae. The analysis gave support for establishing the new tribe Echiochileae for Antiphytum, Echio- chilon and Ogastemma, and for merging the traditionally accepted tribe Eritrichieae with Cyno- glosseae. The ITS region was sequenced for all but one species of Echiochilon and for representa- tives of Antiphytum and Ogastemma. Phylogenetic analysis of Echiochilon revealed that the strongly zygomorphic-flowered species form a paraphyletic group. The morphological data gave results fairly congruent with the ITS phylogeny. Biogeographic interpretations of the ITS and atpB phylogenies indicated a trans-Atlantic dispersal of Antiphytum as the most plausible explanation to the Old/New World disjunction. Analyses using DIVA (Dispersal Vicariance Analysis) of the distributions of the Echiochilon spe- cies indicated an ancestor to Echiochilon with a wide distribution over northern Africa and Arabia to India.