<I>Mahonia Bealei</I>
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

APPROVED PLANT LIST Midtown Alliance Tree Well Adoption Program
APPROVED PLANT LIST Midtown Alliance Tree Well Adoption Program Midtown Alliance launched the Tree Well Adoption program with the primary goal of enriching the experience of Midtown’s workers and residents while encouraging sustainability through the use of low-water, urban tolerant plant species. This list of plants was created to aid individuals and organizations in selecting plant material to plant in their adopted tree wells. This plant list is intended to encourage individual character in the tree wells, rather than restrict creativity in the selection of plants. The plants on the approved list were selected based on the following criteria: • Perennial. All plants listed are perennial, meaning they last for two or more growing seasons. Once established, these plants will require less water to maintain than annuals. • Heat tolerant. Plants in tree wells are exposed to high temperatures caused by vehicles and heat reflected from surrounding buildings, asphalt, and other urban surfaces. They must also be tolerant to high daytime temperatures, typical of Atlanta’s summer months, and cold hardy in the winter months. Atlanta is located in USDA Plant Hardiness Zone 7b/8a. • Water wise. Urban tree wells are surrounded by impervious surfaces and thus, are highly susceptible to periods of drought. Suitable plants must be able to survive periods of low rainfall. • Pollution tolerant. Vehicle exhaust may leave deposits and pollutants on plant foliage, which can kill sensitive plants. • Encourage wildlife. Flowering plants attract insects such as butterflies while others provide food sources for birds and other wildlife. • Grown locally. Many of the plants listed are native to the Atlanta area, and all can be found at local nurseries. -

Shrub List for Brighton 2010
Shrub List For Brighton 2010 Large Shrubs 10’ -20’ Tall by 6’ – 25’ wide Acer ginnala Amur Maple Acer tataricum Tatarian Maple (better than Amur Maple) Acer grandidentatum Bigtooth Maple Amelanchier alnifolia Saskatoon Serviceberry Amelanchier canadensis Shadblow Serviceberry Caragana arborescens Siberian Peashrub Cercocarpus ledifolius Mountain Mahogany Cotoneaster lucidus Peking Cotoneaster Cowania mexicana Quince Bush, Cliffrose Crataefus ambigua Russian Hawthorn Forestiera neomexicana New Mexican Privet Hippophae rhamnoides Sea Buckthorn Juniperus species Juniper Kolkwitzia amabilis Beauty Bush Pinus mugo Mugo Pine species Prunus americana American Plum Prunus virginiana ‘Shubert’ Canada Red Chokecherry Ptelea trifoliata Wafer Ash or Hop tree Quercus gambelii Gambel Oak Rhus typhina Staghorn Sumac Robinia neomexicana New Mexico Locust Sambucus species Elders Shepherdia argentea Buffaloberry Syringa vulgaris Common Lilac Viburnum lantana Wayfaring Tree, Viburnum Medium Size Shrubs >10’ high by >8’ wide Amorpha fruticosa False Indigo Atriplex canescens Fourwing Saltbush Buddleia davidii Butterfly Bush Cercocarpus montanus Mountain Mahogany Chamaebatiaria millefolium Fernbush Chrysothamnus nauseosus Rubber Rabbitbrush Cornus sericea Redtwig Dogwood Cotinus coggygria Smoke Tree Cotoneaster species Cotoneaster Cytisus scoparius ‘Moonlight’ Moonlight Broom Euonymus alatus Burning Bush Forsythia x intermedia Forsythia Hibiscus syriacus Rose-of-Sharon Juniperus species Juniper Ligustrum vulgare Privet Lonicera species Honeysuckle Mahonia aquifolium Oregon Grape Holly Philadelphus species Mockorange Pyracantha coccinea Firethorn Physocarpus opulifolius Common Ninebark Prunus besseyi Western Sand Cherry Pyracantha coccinea species Firethorn Rhamnus frangula Glossy Buckthorn Ribes species Currant Sambucus species Elder Spiraea x vanhouttei Vanhouttei Spirea Symphoricarpos albus Snowberry Syringa meyeri „Palibin‟ Dwarf Korean Lilac Syringa patula „Miss Kim‟ Dwarf Lilac Viburnum species (dozens of different types) Small Size Shrubs > 5’ tall by >6. -

Propagation of Colorado Natives at Little Valley©
224 Combined Proceedings International Plant Propagators’ Society, Volume 58, 2008 Propagation of Colorado Natives at Little Valley © Brian Core Little Valley Wholesale Nursery, 13022 E. 136th Ave., Brighton, Colorado 80601 U.S.A. Email: [email protected] INTRODUCTION Little Valley Wholesale Nursery was established in 1979 with 15 acres of field and container stock and a staff of four people. Over the years, it has expanded to en- compass 141 acres and a peak-season staff of more than 150 people. The plant palette now includes more than 500 taxa of perennials, 275 taxa of shrubs, and over 130 taxa of trees. The area served by Little Valley’s distribution system includes Wyoming, Colorado, New Mexico, and Arizona. Little Valley’s motto “The Rocky Mountain Standard” expresses the commitment to be the best wholesale plant dis- tributor in the western region. Native plants have been an important part of Little Valley’s plant mix for many years. Native plants are ideal for low-input sustainable landscapes. The Denver Metro area receives 8–15 inches of rain per year; in contrast, many eastern cities receive 50 or even 60 inches of annual precipitation. Colorado has a rapidly expanding population that threatens to deplete the water supply within a few decades. Many native plants require no supplemental water when established in the landscape; water that is not wasted on bluegrass lawns is water that can be used for human consumption. Native plants also require less frequent fertilizer and pesticide appli- cations; overuse of fertilizer and pesticide can lead to contamination of groundwater supplies with carcinogens. -

Greg Butler-Bling on the Wing
Bling on the Wing: 12 Months of Flowers for Hummingbirds Greg Butler, BLA, ATA Email: [email protected] Northwest Flower and Garden Festival 2020 Washington species: Anna’s, Rufous, Calliope, Black-Chinned Anna’s: Year-round resident, our largest hummer (4”). Males have bright red/pink gorget, females iridescent green Rufous: Migrate from Southern U.S./Mexico to Alaska. Smaller than Anna’s, about 3-3.5”. Males tawny orange with orange gorget, females green with orange highlights. Recently sighted as far east as Florida and as far north as New Brunswick. Calliope: Primarily found in Eastern WA, occasionally seen on W. slope of Cacades, rare in Puget lowlands. Our smallest hummer, about 2.5-3”. Migrates from Central Mexico as far North as Central BC and Alberta. Males have magenta gorget, females green on back with peachy chest. Black-chinned: primarily found in E Washington, migration pattern similar to Calliope. About same size as Rufous, a bit more slender. Males have black chin/hood with purple gorget, females dull green on back with white chest. Fun facts: Wings can beat up to 80 x per second; diving speeds of up to 50 mph; horizontal speed up to 35 mph; can fly upside down for short distances. Can go from 35 mph to full stop in 6”; can go into state of torpor in cold weather or when hungry. Best eyesight of any bird; amazing pollinators, flowers don’t have to be tubular or red. Can be fiercely territorial (especially Rufous); “hawking” behavior when defending territory, hunting Diet Primarily nectar, insects, and occasionally sap; females have been observed eating ash during nesting season Mating: Males perform impressive courtship dives; females do all of the work of nest building, incubation, and feeding young. -

Alphabetical Lists of the Vascular Plant Families with Their Phylogenetic
Colligo 2 (1) : 3-10 BOTANIQUE Alphabetical lists of the vascular plant families with their phylogenetic classification numbers Listes alphabétiques des familles de plantes vasculaires avec leurs numéros de classement phylogénétique FRÉDÉRIC DANET* *Mairie de Lyon, Espaces verts, Jardin botanique, Herbier, 69205 Lyon cedex 01, France - [email protected] Citation : Danet F., 2019. Alphabetical lists of the vascular plant families with their phylogenetic classification numbers. Colligo, 2(1) : 3- 10. https://perma.cc/2WFD-A2A7 KEY-WORDS Angiosperms family arrangement Summary: This paper provides, for herbarium cura- Gymnosperms Classification tors, the alphabetical lists of the recognized families Pteridophytes APG system in pteridophytes, gymnosperms and angiosperms Ferns PPG system with their phylogenetic classification numbers. Lycophytes phylogeny Herbarium MOTS-CLÉS Angiospermes rangement des familles Résumé : Cet article produit, pour les conservateurs Gymnospermes Classification d’herbier, les listes alphabétiques des familles recon- Ptéridophytes système APG nues pour les ptéridophytes, les gymnospermes et Fougères système PPG les angiospermes avec leurs numéros de classement Lycophytes phylogénie phylogénétique. Herbier Introduction These alphabetical lists have been established for the systems of A.-L de Jussieu, A.-P. de Can- The organization of herbarium collections con- dolle, Bentham & Hooker, etc. that are still used sists in arranging the specimens logically to in the management of historical herbaria find and reclassify them easily in the appro- whose original classification is voluntarily pre- priate storage units. In the vascular plant col- served. lections, commonly used methods are systema- Recent classification systems based on molecu- tic classification, alphabetical classification, or lar phylogenies have developed, and herbaria combinations of both. -

Dipterists Forum
BULLETIN OF THE Dipterists Forum Bulletin No. 76 Autumn 2013 Affiliated to the British Entomological and Natural History Society Bulletin No. 76 Autumn 2013 ISSN 1358-5029 Editorial panel Bulletin Editor Darwyn Sumner Assistant Editor Judy Webb Dipterists Forum Officers Chairman Martin Drake Vice Chairman Stuart Ball Secretary John Kramer Meetings Treasurer Howard Bentley Please use the Booking Form included in this Bulletin or downloaded from our Membership Sec. John Showers website Field Meetings Sec. Roger Morris Field Meetings Indoor Meetings Sec. Duncan Sivell Roger Morris 7 Vine Street, Stamford, Lincolnshire PE9 1QE Publicity Officer Erica McAlister [email protected] Conservation Officer Rob Wolton Workshops & Indoor Meetings Organiser Duncan Sivell Ordinary Members Natural History Museum, Cromwell Road, London, SW7 5BD [email protected] Chris Spilling, Malcolm Smart, Mick Parker Nathan Medd, John Ismay, vacancy Bulletin contributions Unelected Members Please refer to guide notes in this Bulletin for details of how to contribute and send your material to both of the following: Dipterists Digest Editor Peter Chandler Dipterists Bulletin Editor Darwyn Sumner Secretary 122, Link Road, Anstey, Charnwood, Leicestershire LE7 7BX. John Kramer Tel. 0116 212 5075 31 Ash Tree Road, Oadby, Leicester, Leicestershire, LE2 5TE. [email protected] [email protected] Assistant Editor Treasurer Judy Webb Howard Bentley 2 Dorchester Court, Blenheim Road, Kidlington, Oxon. OX5 2JT. 37, Biddenden Close, Bearsted, Maidstone, Kent. ME15 8JP Tel. 01865 377487 Tel. 01622 739452 [email protected] [email protected] Conservation Dipterists Digest contributions Robert Wolton Locks Park Farm, Hatherleigh, Oakhampton, Devon EX20 3LZ Dipterists Digest Editor Tel. -

Vascular Plant and Vertebrate Inventory of Montezuma Castle National Monument Vascular Plant and Vertebrate Inventory of Montezuma Castle National Monument
Schmidt, Drost, Halvorson In Cooperation with the University of Arizona, School of Natural Resources Vascular Plant and Vertebrate Inventory of Montezuma Castle National Monument Vascular Plant and Vertebrate Inventory of Montezuma Castle National Monument Plant and Vertebrate Vascular U.S. Geological Survey Southwest Biological Science Center 2255 N. Gemini Drive Flagstaff, AZ 86001 Open-File Report 2006-1163 Southwest Biological Science Center Open-File Report 2006-1163 November 2006 U.S. Department of the Interior U.S. Geological Survey National Park Service In cooperation with the University of Arizona, School of Natural Resources Vascular Plant and Vertebrate Inventory of Montezuma Castle National Monument By Cecilia A. Schmidt, Charles A. Drost, and William L. Halvorson Open-File Report 2006-1163 November, 2006 USGS Southwest Biological Science Center Sonoran Desert Research Station University of Arizona U.S. Department of the Interior School of Natural Resources U.S. Geological Survey 125 Biological Sciences East National Park Service Tucson, Arizona 85721 U.S. Department of the Interior Dirk Kempthorne, Secretary U.S. Geological Survey Mark Myers, Director U.S. Geological Survey, Reston, Virginia: 2006 Note: This document contains information of a preliminary nature and was prepared primarily for internal use in the U.S. Geological Survey. This information is NOT intended for use in open literature prior to publication by the investigators named unless permission is obtained in writing from the investigators named and from the Station Leader. Suggested Citation Schmidt, C. A., C. A. Drost, and W. L. Halvorson 2006. Vascular Plant and Vertebrate Inventory of Montezuma Castle National Monument. USGS Open-File Report 2006-1163. -

Native Or Suitable Plants City of Mccall
Native or Suitable Plants City of McCall The following list of plants is presented to assist the developer, business owner, or homeowner in selecting plants for landscaping. The list is by no means complete, but is a recommended selection of plants which are either native or have been successfully introduced to our area. Successful landscaping, however, requires much more than just the selection of plants. Unless you have some experience, it is suggested than you employ the services of a trained or otherwise experienced landscaper, arborist, or forester. For best results it is recommended that careful consideration be made in purchasing the plants from the local nurseries (i.e. Cascade, McCall, and New Meadows). Plants brought in from the Treasure Valley may not survive our local weather conditions, microsites, and higher elevations. Timing can also be a serious consideration as the plants may have already broken dormancy and can be damaged by our late frosts. Appendix B SELECTED IDAHO NATIVE PLANTS SUITABLE FOR VALLEY COUNTY GROWING CONDITIONS Trees & Shrubs Acer circinatum (Vine Maple). Shrub or small tree 15-20' tall, Pacific Northwest native. Bright scarlet-orange fall foliage. Excellent ornamental. Alnus incana (Mountain Alder). A large shrub, useful for mid to high elevation riparian plantings. Good plant for stream bank shelter and stabilization. Nitrogen fixing root system. Alnus sinuata (Sitka Alder). A shrub, 6-1 5' tall. Grows well on moist slopes or stream banks. Excellent shrub for erosion control and riparian restoration. Nitrogen fixing root system. Amelanchier alnifolia (Serviceberry). One of the earlier shrubs to blossom out in the spring. -

Diptera: Tephritidae)
ANNALS OF THE UPPER SILESIAN MUSEUM IN BYTOM ENTOMOLOGY Vol. 28 (online 008): 1–9 ISSN 0867-1966, eISSN 2544-039X (online) Bytom, 17.12.2019 ANDRZEJ PALACZYK1 , ANNA KLASA2, ANDRZEJ SZLACHETKA3 First record in Poland and remarks on the origin of the northern populations of Goniglossum wiedemanni MEIGEN, 1826 (Diptera: Tephritidae) http://doi.org/10.5281/zenodo.3580897 1 Institute of Systematics and Evolution of Animals, Polish Academy of Sciences, Sławkowska 17, 31–016 Kraków, Poland, e-mail: [email protected] 2 Ojców National Park, 32–045 Sułoszowa, Ojców 9, e-mail: [email protected] 3 Parszowice 81, 59–330 Ścinawa, e-mail: [email protected] Abstract: The fruit fly Goniglossum wiedemanni has been recorded from Poland for the first time. Found in a single locality (Parszowice) in Lower Silesia, this species was recorded in a garden on Bryonia alba. Notes on the identification, biology and remarks on the general distribution and origin of the northern populations of this species are given. Colour photographs of the habitus and live specimens are also provided. Key words: Goniglossum wiedemanni, Carpomyini, species new for Poland, Lower Silesia, general distribution, Bryonia alba. INTRODUCTION Species from the family Tephritidae, the larvae of which develop in fruit, belong to the subfamilies Dacinae and Trypetinae. They occur most numerously in regions with a tropical or subtropical climate, where they pose a serious economic problem: in some areas they give rise to crop losses worth many millions of dollars. In central Europe, there are only a few species whose larvae feed on fruit; they belong exclusively to the tribes Carpomyini and Trypetini from the subfamily Trypetinae. -
Show Activity
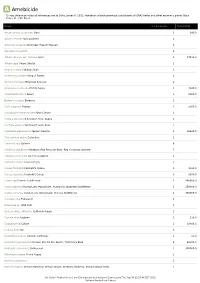
A Amebicide *Unless otherwise noted all references are to Duke, James A. 1992. Handbook of phytochemical constituents of GRAS herbs and other economic plants. Boca Raton, FL. CRC Press. Plant # Chemicals Total PPM Abelmoschus esculentus Okra 1 140.0 Adonis vernalis Spring Adonis 1 Ailanthus altissima Stinktree; Tree Of Heaven 3 Alangium lamarckii 2 Allium sativum var. sativum Garlic 2 57531.0 Allium cepa Onion; Shallot 1 Andira inermis Cabbage Bark 1 Anemone pulsatilla Pasque Flower 1 Annona montana Mountain Soursop 1 Argemone mexicana Prickly Poppy 1 1640.0 Azadirachta indica Neem 1 8000.0 Berberis vulgaris Barberry 1 Carica papaya Papaya 1 5500.0 Caulophyllum thalictroides Blue Cohosh 1 Ceiba pentandra Silk-Cotton Tree; Kapok 1 Centella asiatica Pennywort; Gotu Kola 1 Cephaelis ipecacuanha Ipecac; Raicilla 3 96900.0 Chelidonium majus Celandine 1 Cinchona spp Quinine 6 Cinchona pubescens Redbark; Red Peruvian-Bark; Red Cinchona; Quinine 7 Cinchona officinalis Quinine; Lojabark 4 Clematis vitalba Traveler's Joy 1 Coleus forskohlii Forskohl's Coleus 1 9260.0 Coleus barbatus Forskohl's Coleus 1 9260.0 Coptis spp Generic Goldthread 2 360000.0 Coptis japonica Huang-Lian; Huang-Lien; Huang-Lia; Japanese Goldthread 1 280000.0 Coptis chinensis Huang-Lien; Huang-Lian; Chinese Goldthread 1 360000.0 Corydalis spp Fumewort 1 Dioscorea sp. Wild Yam 1 Eschscholzia californica California Poppy 1 Glycine max Soybean 1 120.0 Gossypium sp Cotton 1 19000.0 Hedera helix Ivy 1 Helianthus annuus Girasol; Sunflower 1 20.0 Holarrhena pubescens Conessi; Zhi Xie Mu; Kurchi; Tellicherry Bark 2 26000.0 Hydrastis canadensis Goldenseal 2 300000.0 Macleaya cordata Plume Poppy 1 Mahonia siamensis 1 Mahonia repens Oregon Barberry; Oregon Grape; Creeping Barberry; Oregon Grape-Holly 1 Dr. -

OSU Gardening with Oregon Native Plants
GARDENING WITH OREGON NATIVE PLANTS WEST OF THE CASCADES EC 1577 • Reprinted March 2008 CONTENTS Benefi ts of growing native plants .......................................................................................................................1 Plant selection ....................................................................................................................................................2 Establishment and care ......................................................................................................................................3 Plant combinations ............................................................................................................................................5 Resources ............................................................................................................................................................5 Recommended native plants for home gardens in western Oregon .................................................................8 Trees ...........................................................................................................................................................9 Shrubs ......................................................................................................................................................12 Groundcovers ...........................................................................................................................................19 Herbaceous perennials and ferns ............................................................................................................21 -
Fire-Resistant Plants for Oregon Home Landscapes
FFire-ire-RResistantesistant PlantsPlants forfor OregonOregon HomeHome LandscapesLandscapes Suggesting specific types of vegetation that may reduce your risk from wildfire. Stephen Fitzgerald Area Extension Forester and Associate Professor Amy Jo Waldo Area Extension Horticulture Agent and Assistant Professor OSU Extension Service 1421 S. Hwy 97, Redmond, OR 97756 Introduction Oregon has many wildfire prone areas. In these places, fires are a natural part of the changing landscape. As homes are built in these areas, special precautions must be taken by the homeowner to pro- tect their property. Installation of fire- resistive roofing is critical to preventing firebrands from igniting the home from a roof fire. Well maintained fire-resistant vegetation and irrigated landscape is also critical within close proximity of a home. These actions DO NOT insure that your home will survive a wildfire, but they provide for a good chance of structural survival. Implementation of FireFree [www.firefree.org] and FireWise [www.firewise.org] activities can also significantly improve chances of a home surviving a wildfire. Fire-resistant vegetation. When landscaping around a home, most homeowners are interested in creating a landscape that is aesthetically pleasing, compliments their home, and has varia- tions in color, texture, flowers, and foliage. If your home is located in or adjacent to forests or rangeland, you should also consider the flammability of plants within your home landscape. Flammable plant material in your land- scape can increase the fire-risk around your home. The 1991 Oakland Hills Fire in California is a prime example of how flam- mable plant material (Eucalyptus trees) can act as fuel and contribute to the inten- sity of a wildfire.