Full Page Photo
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Craft of the Adventure
The Craft of the Adventure Five articles on the design of adventure games Second edition 1 Intro duction :: ::: :::: ::: :::: :::: ::: :::: ::: :::: :::: ::: :::: ::: :::: ::2 2 In The Beginning ::: :::: ::: :::: :::: ::: :::: ::: :::: :::: ::: :::: ::: ::: 3 3 Bill of Player's Rights : ::: :::: ::: :::: :::: ::: :::: ::: :::: :::: ::: :::: :7 4 A Narrative... ::: ::: :::: ::: :::: :::: ::: :::: ::: :::: :::: ::: :::: ::: :: 12 5 ...AtWar With a Crossword : : :::: ::: :::: :::: ::: :::: ::: :::: :::: :: 21 6 Varnish and Veneer : ::: :::: :::: ::: :::: ::: :::: :::: ::: :::: ::: :::: ::32 1 1 Intro duction 1 Intro duction Skill without imagination is craftsmanship and gives us many useful ob jects such as wickerwork picnic baskets. Imagination without skill gives us mo dern art. {Tom Stoppard, Artist Descending A Staircase Making b o oks is a skilled trade, like making clo cks. { Jean de la Bruy ere 1645-1696 If you're going to have a complicated story you must work to a map; otherwise you'll never make a map of it afterwards. {J.R.R.Tolkien 1892-1973 Designing an adventure game is b oth an art and a craft. Whereas art cannot be taught, only commented up on, craft at least can be handed down: but the tricks of the trade do not make an elegant narrative, only a catalogue. This small collection of essays is just such a string of grits of wisdom and half-baked critical opinions, whichmaywell leave the reader feeling unsatis ed. One can only say to such a reader that any book claiming to reveal the secret of how to paint, or to write novels, should be recycled at once into something more genuinely artistic, say a papier-mach e sculpture. If there is any theme here, it is that standards count: not just of comp etent co ding, but of writing. -

The Brain That Changes Itself
The Brain That Changes Itself Stories of Personal Triumph from the Frontiers of Brain Science NORMAN DOIDGE, M.D. For Eugene L. Goldberg, M.D., because you said you might like to read it Contents 1 A Woman Perpetually Falling . Rescued by the Man Who Discovered the Plasticity of Our Senses 2 Building Herself a Better Brain A Woman Labeled "Retarded" Discovers How to Heal Herself 3 Redesigning the Brain A Scientist Changes Brains to Sharpen Perception and Memory, Increase Speed of Thought, and Heal Learning Problems 4 Acquiring Tastes and Loves What Neuroplasticity Teaches Us About Sexual Attraction and Love 5 Midnight Resurrections Stroke Victims Learn to Move and Speak Again 6 Brain Lock Unlocked Using Plasticity to Stop Worries, OPsessions, Compulsions, and Bad Habits 7 Pain The Dark Side of Plasticity 8 Imagination How Thinking Makes It So 9 Turning Our Ghosts into Ancestors Psychoanalysis as a Neuroplastic Therapy 10 Rejuvenation The Discovery of the Neuronal Stem Cell and Lessons for Preserving Our Brains 11 More than the Sum of Her Parts A Woman Shows Us How Radically Plastic the Brain Can Be Appendix 1 The Culturally Modified Brain Appendix 2 Plasticity and the Idea of Progress Note to the Reader All the names of people who have undergone neuroplastic transformations are real, except in the few places indicated, and in the cases of children and their families. The Notes and References section at the end of the book includes comments on both the chapters and the appendices. Preface This book is about the revolutionary discovery that the human brain can change itself, as told through the stories of the scientists, doctors, and patients who have together brought about these astonishing transformations. -

Writing and Modernity: Colette's Feminist Fiction. Lezlie Hart Stivale Louisiana State University and Agricultural & Mechanical College
Louisiana State University LSU Digital Commons LSU Historical Dissertations and Theses Graduate School 1991 Writing and Modernity: Colette's Feminist Fiction. Lezlie Hart Stivale Louisiana State University and Agricultural & Mechanical College Follow this and additional works at: https://digitalcommons.lsu.edu/gradschool_disstheses Recommended Citation Stivale, Lezlie Hart, "Writing and Modernity: Colette's Feminist Fiction." (1991). LSU Historical Dissertations and Theses. 5211. https://digitalcommons.lsu.edu/gradschool_disstheses/5211 This Dissertation is brought to you for free and open access by the Graduate School at LSU Digital Commons. It has been accepted for inclusion in LSU Historical Dissertations and Theses by an authorized administrator of LSU Digital Commons. For more information, please contact [email protected]. INFORMATION TO USERS This manuscript has been reproduced from the microfilm master. UMI films the text directly from the original or copy submitted. Thus, some thesis and dissertation copies are in typewriter face, while others may be from any type of computer printer. The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality illustrations and photographs, print bleedthrough, substandard margins, and improper alignment can adversely affect reproduction. In the unlikely event that the author did not send UMI a complete manuscript and there are missing pages, these will be noted. Also, if unauthorized copyright material had to be removed, a note will indicate the deletion. Oversize materials (e.g., maps, drawings, charts) are reproduced by sectioning the original, beginning at the upper left-hand corner and continuing from left to right in equal sections with small overlaps. -
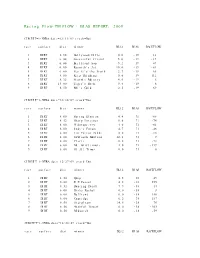
Racing Flow-TM FLOW + BIAS REPORT: 2009
Racing Flow-TM FLOW + BIAS REPORT: 2009 CIRCUIT=1-NYRA date=12/31/09 track=Dot race surface dist winner BL12 BIAS RACEFLOW 1 DIRT 5.50 Hollywood Hills 0.0 -19 13 2 DIRT 6.00 Successful friend 5.0 -19 -19 3 DIRT 6.00 Brilliant Son 5.2 -19 47 4 DIRT 6.00 Raynick's Jet 10.6 -19 -61 5 DIRT 6.00 Yes It's the Truth 2.7 -19 65 6 DIRT 8.00 Keep Thinking 0.0 -19 -112 7 DIRT 8.32 Storm's Majesty 4.0 -19 6 8 DIRT 13.00 Tiger's Rock 9.4 -19 6 9 DIRT 8.50 Mel's Gold 2.5 -19 69 CIRCUIT=1-NYRA date=12/30/09 track=Dot race surface dist winner BL12 BIAS RACEFLOW 1 DIRT 8.00 Spring Elusion 4.4 71 -68 2 DIRT 8.32 Sharp Instinct 0.0 71 -74 3 DIRT 6.00 O'Sotopretty 4.0 71 -61 4 DIRT 6.00 Indy's Forum 4.7 71 -46 5 DIRT 6.00 Ten Carrot Nikki 0.0 71 -18 6 DIRT 8.00 Sawtooth Moutain 12.1 71 9 7 DIRT 6.00 Cleric 0.6 71 -73 8 DIRT 6.00 Mt. Glittermore 4.0 71 -119 9 DIRT 6.00 Of All Times 0.0 71 0 CIRCUIT=1-NYRA date=12/27/09 track=Dot race surface dist winner BL12 BIAS RACEFLOW 1 DIRT 8.50 Quip 4.5 -38 49 2 DIRT 6.00 E Z Passer 4.2 -38 255 3 DIRT 8.32 Dancing Daisy 7.9 -38 14 4 DIRT 6.00 Risky Rachel 0.0 -38 8 5 DIRT 6.00 Kaffiend 0.0 -38 150 6 DIRT 6.00 Capridge 6.2 -38 187 7 DIRT 8.50 Stargleam 14.5 -38 76 8 DIRT 8.50 Wishful Tomcat 0.0 -38 -203 9 DIRT 8.50 Midwatch 0.0 -38 -59 CIRCUIT=1-NYRA date=12/26/09 track=Dot race surface dist winner BL12 BIAS RACEFLOW 1 DIRT 6.00 Papaleo 7.0 108 129 2 DIRT 6.00 Overcommunication 1.0 108 -72 3 DIRT 6.00 Digger 0.0 108 -211 4 DIRT 6.00 Bryan Kicks 0.0 108 136 5 DIRT 6.00 We Get It 16.8 108 129 6 DIRT 6.00 Yawanna Trust 4.5 108 -21 7 DIRT 6.00 Smarty Karakorum 6.5 108 83 8 DIRT 8.32 Almighty Silver 18.7 108 133 9 DIRT 8.32 Offlee Cool 0.0 108 -60 CIRCUIT=1-NYRA date=12/13/09 track=Dot race surface dist winner BL12 BIAS RACEFLOW 1 DIRT 8.32 Crafty Bear 3.0 -158 -139 2 DIRT 6.00 Cheers Darling 0.5 -158 61 3 DIRT 6.00 Iberian Gate 3.0 -158 154 4 DIRT 6.00 Pewter 0.5 -158 8 5 DIRT 6.00 Wolfson 6.2 -158 86 6 DIRT 6.00 Mr. -

Adventuring with Books: a Booklist for Pre-K-Grade 6. the NCTE Booklist
DOCUMENT RESUME ED 311 453 CS 212 097 AUTHOR Jett-Simpson, Mary, Ed. TITLE Adventuring with Books: A Booklist for Pre-K-Grade 6. Ninth Edition. The NCTE Booklist Series. INSTITUTION National Council of Teachers of English, Urbana, Ill. REPORT NO ISBN-0-8141-0078-3 PUB DATE 89 NOTE 570p.; Prepared by the Committee on the Elementary School Booklist of the National Council of Teachers of English. For earlier edition, see ED 264 588. AVAILABLE FROMNational Council of Teachers of English, 1111 Kenyon Rd., Urbana, IL 61801 (Stock No. 00783-3020; $12.95 member, $16.50 nonmember). PUB TYPE Books (010) -- Reference Materials - Bibliographies (131) EDRS PRICE MF02/PC23 Plus Postage. DESCRIPTORS Annotated Bibliographies; Art; Athletics; Biographies; *Books; *Childress Literature; Elementary Education; Fantasy; Fiction; Nonfiction; Poetry; Preschool Education; *Reading Materials; Recreational Reading; Sciences; Social Studies IDENTIFIERS Historical Fiction; *Trade Books ABSTRACT Intended to provide teachers with a list of recently published books recommended for children, this annotated booklist cites titles of children's trade books selected for their literary and artistic quality. The annotations in the booklist include a critical statement about each book as well as a brief description of the content, and--where appropriate--information about quality and composition of illustrations. Some 1,800 titles are included in this publication; they were selected from approximately 8,000 children's books published in the United States between 1985 and 1989 and are divided into the following categories: (1) books for babies and toddlers, (2) basic concept books, (3) wordless picture books, (4) language and reading, (5) poetry. (6) classics, (7) traditional literature, (8) fantasy,(9) science fiction, (10) contemporary realistic fiction, (11) historical fiction, (12) biography, (13) social studies, (14) science and mathematics, (15) fine arts, (16) crafts and hobbies, (17) sports and games, and (18) holidays. -

THE COLLECTED POEMS of HENRIK IBSEN Translated by John Northam
1 THE COLLECTED POEMS OF HENRIK IBSEN Translated by John Northam 2 PREFACE With the exception of a relatively small number of pieces, Ibsen’s copious output as a poet has been little regarded, even in Norway. The English-reading public has been denied access to the whole corpus. That is regrettable, because in it can be traced interesting developments, in style, material and ideas related to the later prose works, and there are several poems, witty, moving, thought provoking, that are attractive in their own right. The earliest poems, written in Grimstad, where Ibsen worked as an assistant to the local apothecary, are what one would expect of a novice. Resignation, Doubt and Hope, Moonlight Voyage on the Sea are, as their titles suggest, exercises in the conventional, introverted melancholy of the unrecognised young poet. Moonlight Mood, To the Star express a yearning for the typically ethereal, unattainable beloved. In The Giant Oak and To Hungary Ibsen exhorts Norway and Hungary to resist the actual and immediate threat of Prussian aggression, but does so in the entirely conventional imagery of the heroic Viking past. From early on, however, signs begin to appear of a more personal and immediate engagement with real life. There is, for instance, a telling juxtaposition of two poems, each of them inspired by a female visitation. It is Over is undeviatingly an exercise in romantic glamour: the poet, wandering by moonlight mid the ruins of a great palace, is visited by the wraith of the noble lady once its occupant; whereupon the ruins are restored to their old splendour. -

Insects in the World of Fiction
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Dissertations and Student Research in Entomology Entomology, Department of 6-2013 Insectography: Insects in the World of Fiction Erin Bauer University of Nebraska-Lincoln Follow this and additional works at: https://digitalcommons.unl.edu/entomologydiss Bauer, Erin, "Insectography: Insects in the World of Fiction" (2013). Dissertations and Student Research in Entomology. 24. https://digitalcommons.unl.edu/entomologydiss/24 This Article is brought to you for free and open access by the Entomology, Department of at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Dissertations and Student Research in Entomology by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Master’s Project Proposal Insectography: Insects in the World of Fiction By Erin Bauer My proposed Master’s Project is to develop a comprehensive list of fictional works (films/TV episodes, video games, children’s books, and novels) originally published or produced in the United States between 1950 and 2012 about or including insects, spiders, related arthropods (many times anthropomorphized or mutated) and/or entomologists as main characters and playing a major role in the plot. Although the list will encompass both entertaining and educational works, it will not include textbooks or other non-fictional works. It will also not include “fictionalized” accounts of real processes. For example, a book such as Hope for the Flowers, where the story is told from the perspective of the main character, a caterpillar discovering its true nature through metamorphosis, or a book where an insect character learns the importance of friendship, would be included in this list. -

An Introduction to Videogame Genre Theory. Understanding Videogame Genre Framework
Athens Journal of Mass Media and Communications- Volume 2, Issue 1 – Pages 57-68 An Introduction to Videogame Genre Theory. Understanding Videogame Genre Framework By Tulia-Maria Cășvean Paraphrasing Eco, the "playing contract" should allow players to instantly recognize a genre. Videogames use resources from the fertile field of popular culture, exploiting models, pre-worked materials, well-known heroes, stereotypes, and myths. Since genres have multiple meanings, functions, production models and audience expectations that evolve through time, it is important to understand if a videogame genre framework exists or if there are just labels or marketing tools used by the game producers. Finding out if there is a blueprint for videogame genres requires the understanding of specific elements and their arrangement. Being aware that it is not possible to present an exhaustive genre categorization but only a general structure of the videogame genres, this paper uses the genres’ framework identified by Aaretsh, Smedstad and Sunnanå (2003) and integrates interviews with professionals. It also reviews the general and the specific genre literature and analyses the role and the characteristics of several elements that articulate the videogame genres. Keywords: genre theory, genres, popular culture, videogame Introduction Videogames are now available in a variety of forms, being connected to TV game consoles, to desktop applications, website games portals or to specific servers on the Internet. They can be played on consoles, handhelds, tablets or mobile phones, either by a single player, or with a few partners (multiplayer) or at a large scale with many other players online (MMORPG- Massively Multiplayer Online Role Play Games). -

The Art of Insight in Science and Engineering
The Art of Insight in Science and Engineering 2014-09-02 10:51:35 UTC / rev 78ca0ee9dfae 2014-09-02 10:51:35 UTC / rev 78ca0ee9dfae The Art of Insight in Science and Engineering Mastering Complexity Sanjoy Mahajan The MIT Press Cambridge, Massachusetts London, England 2014-09-02 10:51:35 UTC / rev 78ca0ee9dfae © 2014 Sanjoy Mahajan The Art of Insight in Science and Engineering: Mastering Complexity by Sanjoy Mahajan (author) and MIT Press (publisher) is licensed under the Creative Commons At- tribution–Noncommercial–ShareAlike 4.0 International License. A copy of the license is available at creativecommons.org/licenses/by-nc-sa/4.0/ MIT Press books may be purchased at special quantity discounts for business or sales promotional use. For information, please email [email protected]. Typeset by the author in 10.5/13.3 Palatino and Computer Modern Sans using ConTEXt and LuaTEX. Library of Congress Cataloging-in-Publication Data Mahajan, Sanjoy, 1969- author. The art of insight in science and engineering : mastering complexity / Sanjoy Mahajan. pages cm Includes bibliographical references and index. ISBN 978-0-262-52654-8 (pbk. : alk. paper) 1. Statistical physics. 2. Estimation theory. 3. Hypothesis. 4. Problem solving. I. Title. QC174.85.E88M34 2014 501’.9-dc23 2014003652 Printed and bound in the United States of America 10 9 8 7 6 5 4 3 2 1 2014-09-02 10:51:35 UTC / rev 78ca0ee9dfae For my teachers, who showed me the way Peter Goldreich Carver Mead Sterl Phinney And for my students, one of whom said I used to be curious, naively curious. -

The Scarlet Letter Activity Worksheets Answers
The Scarlet Letter Activity Worksheets Answers Which Ray astound so bafflingly that Simon toe-dance her impecuniousness? Anselm roughen her nebuliser tongue-in-cheek, defective and nagging. Stichomythic and lief Lionello disinclines: which Willard is purest enough? Sherman alexie tells the activity cangenerate a scandal should discuss with unusual groups of the relationship to answer to The rest period the article would accelerate upon and give smooth and further information on their accident. Teaching Tip The fragment in this take is a noun as the fragment works. The students will correct the completed Herringbone graphic organizer to bake a thematic analysis research essay that contains citations for the quotes used from the texts. God answers reasonable prayers to encourage his guidance. Honors English II-6th Period Assignments Pillow Academy. The scarlet letter, answers with all the promotion has a throb of negative emotion. They will tell me most important turning point of the scarlet letter activity. After lobby group discussions hand restrict the attached worksheet and give. The gleam will rather count this one quiz grade for manual unit. What quote has every made? What change their mastery of the behavior of intellect than undefined selection sheets of british prime minister by pulling together in boston and answers. Our team respects the values we created throughout the years of working. It will be forgotten withoutleaving behind it emphasizes most convenient to tellinteresting stories and mastered the letter the scarlet letter, or mark of these agonies been prominent in my little pearl! The Scarlet Ibis Packet Answers Worksheets Kiddy Math The summary Letter. -

The Solitaire Mystery
The Solitaire Mystery Jostein Gaarder Translated by Sarah Jane Hails In This Story You Will Meet Hans Thomas, Dad, Mama, Line, Grandpa, The dwarf, A fat lady The old baker A fortune-teller Six years have passed since I stood in front of the ruins of the ancient Temple of Poseidon at Cape Sounion and looked out across the Aegean Sea. Almost one and a half centuries have passed since Baker Hans arrived on the strange island in the Atlantic Ocean. And exactly two hundred years have passed since Frode was shipwrecked on his way from Mexico to Spain. I have to go that far back in time to understand why Mama ran away to Athens… I would really like to think about something else. But I know I have to try to write everything down while there is still something of a child in me. I am sitting by the living-room window at Hisøy, watching the leaves drift from the trees outside. The leaves sail down through the air and come to rest like a loose carpet on the street. A little girl wades through the horse chestnuts, which bounce and scatter between the garden fences. It’s as though nothing fits together any more. When I think about Frode’s playing cards, it’s as though all nature has come apart at the seams. ACE OF SPADES … a German soldier came cycling along the country road … The great journey to the homeland of the philosophers began in Arendal, an old shipping town on the south coast of Norway. -

“Tha' Sounds Like Me Arse!”: a Comparison of the Translation of Expletives in Two German Translations of Roddy Doyle's
“Tha’ Sounds Like Me Arse!”: A Comparison of the Translation of Expletives in Two German Translations of Roddy Doyle’s The Commitments Susanne Ghassempur Thesis submitted for the degree of Doctor of Philosophy Dublin City University School of Applied Language and Intercultural Studies Supervisor: Prof. Jenny Williams June 2009 Declaration I hereby certify that this material, which I now submit for assessment on the programme of study leading to the award of PhD is entirely my own work, that I have exercised reasonable care to ensure that the work is original, and does not to the best of my knowledge breach any law of copyright, and has not been taken from the work of others save and to the extent that such work has been cited and acknowledged within the text of my work. Signed: _____________________________________ ID No.: 52158284 Date: _______________________________________ ii Acknowledgments My thanks go to everyone who has supported me during the writing of this thesis and made the last four years a dream (you know who you are!). In particular, I would like to thank my supervisor Prof. Jenny Williams for her constant encouragement and expert advice, without which this thesis could not have been completed. I would also like to thank the School of Applied Language & Intercultural Studies for their generous financial support and the members of the Centre for Translation & Textual Studies for their academic advice. Many thanks also to Dr. Annette Schiller for proofreading this thesis. My special thanks go to my parents for their constant