Cadmium Tolerance in a Soil Arthropod a Model of Real-Time Microevolution
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Changing of the Guard in White Grub Control Insecticides
A PRACTICAL RESEARCH DIGEST FOR TURF MANAGERS Volume 10, Issue 6 • June 2001 ¡TURFGRASS PEST CONTROL IN THIS ISSUE • The changing of the The Changing of the guard in white grub Guard in White Grub control insecticides 1 Organophosphate/ Control Insecticides carbamate update New product information By Kevin Mathias Natural control influence of insecticides combination of federal regulatory rulings and economic decisions by insecticide Multiple targeting manufacturers has dramatically changed the landscape of white grub insecticides A and control strategies. At the beginning of the 1990's white grub control insecti- • Site analysis for golf cides consisted mainly of organophosphate and carbamate based chemistries with only a course development 7 few biorational products available (Table 1). As a group, the organophosphate and car- Climate bamate insecticides, have a relatively short residual activity and are highly efficacious when used in curative control programs. Topography Optimum results are attained if the products are applied in mid to late August or into September, as white grub damage is first noticed and Drainage patterns when the grubs are young and relatively small. Optimum results are Water availability As we enter the new millennium many of the cura- attained if the tive control products have been replaced by a group of Soils and geology products are applied new insecticides. These insecticides, Merit and Mach 2, offer greater applicator safety, have less adverse effect Environmental issues in mid to late August on the environment, provide a longer window of appli- Wetlands or into September, as cation due to their extended soil residual activities, have minimal impact on beneficial predators, and pro- Water quality white grub damage vide excellent control (+90%) of white grubs. -

Springtails in Sugarbeet: Identification, Biology, And
E-1205 SpringtailsSpringtails in Sugarbeet: Figure 1. Adult springtail (40x magnification). Identification, Biology, and Management Mark A. Boetel, Research and Extension Entomologist Robert J. Dregseth, Research Specialist Introduction Mohamed F. R. Khan, Extension Sugarbeet Specialist Springtails are tiny, wingless, primitive animals commonly placed into the insect order Collembola. They are so unusual that some experts classify them as non-insects. Springtails Description are one of the most abundant and diverse animal groups on Springtails can vary in color from white to yellow, earth with over 6,000 described species and an estimated orange, metallic green, lavender, gray, or red. A tiny tube eight times as many remaining to be identified. on the abdomen, the collophore, is common to all spring- The springtails have worldwide distribution and tails. The collophore is mostly needed for maintaining occupy a diverse habitat range that includes soil, algae, old optimal water balance but also functions in some species as snowbanks, beaches, caves, cisterns, vacant bird nests, a sticky appendage for adhering to surfaces. The name tropical rain forest canopies, tidal pools, deserts, the “springtail” refers to an unusual forked organ, the furcula, surfaces of freshwater ponds and streams, and even the that arises near the posterior end of some species. It enables frozen terrain of Antarctica. However, they are most them to jump when disturbed. The furcula is usually folded abundant in warm, moist environments. Economically forward along the underside of the body and held in place important species in North Dakota and Minnesota with a clasp called the tenaculum. To jump, the springtail sugarbeet fields are the soil-inhabiting springtails. -

13 Index of Common Names
Index of Common Names BITING/STINGING/VENOMOUS PESTS Bees ………………………………………………………… 6 Honey bee…………………………………………………6 Africanized bees……………………………………….. 7 Bumblebee…………………………………………………….9 Carpenter bee……………………………………………….9 Digger bee………………………………………………………11 Leaf cutter bee………………………………………………….12 Sweat bee………………………………………………………..13 Wasps…………………………………………………………….14 Tarantula hawk…………………………………………………14 Yellowjacket……………………………………………………….15 Aerial yellowjacket ……………………………………………….15,16 Common yellowjacket ……………………………………………….15 German yellowjacket……………………………………………….15 Western yellowjacket……………………………………………….15 Paper wasp………………………………………………. 18 Brown paper wasp……………………………………………….18 Common paper wasp ……………………………………………….18 European paper wasp……………………………………………….18 Navajo paper wasp……………………………………………….18,19 Yellow paper wasp……………………………………………….18,19 Western paper wasp……………………………………………….18 Mud dauber………………………………………………. 20 Black and blue mud dauber……………………………..………………….20 Black and yellow mud dauber……………………………………………….20 Organ pipe mud dauber……………………………………………….20,21 Velvet ant……………………………………………………21 Scorpions………………………………………………………23 Arizona bark scorpion……………………………………………….23 Giant hairy scorpion ……………………………………………….24 Striped-tail scorpion……………………………………………….25 Yellow ground scorpion……………………………………………….25 Spiders…………………………………………………………..26 Cellar spider……………………………………………….2 6 Recluse spider……………………………………………….27 Tarantula…………………………………………………. 28 Widow spider……………………………………………….29 Scorpion/spider look-alikes……………………………………………….30 Pseudoscorpion……………………………………………….30 Solifugid/wind -

Observations on the Springtail Leaping Organ and Jumping Mechanism Worked by a Spring
Observations on the Springtail Leaping Organ and Jumping Mechanism Worked by a Spring Seiichi Sudo a*, Masahiro Shiono a, Toshiya Kainuma a, Atsushi Shirai b, and Toshiyuki Hayase b a Faculty of Systems Science and Technology, Akita Prefectural University, Japan b Institute of Fluid Science, Tohoku University, Japan Abstract— This paper is concerned with a small In this paper, the springtail leaping organ was jumping mechanism. Microscopic observations of observed with confocal laser scanning microscopy. A springtail leaping organs were conducted using a jumping mechanism using a spring and small confocal leaser scanning microscope. A simple electromagnet was produced based on the observations springtail mechanism using a spring and small of leaping organ and the jumping analysis of the electromagnet was produced based on the observations springtail. of leaping organ and the jumping analysis of the globular springtail. Jumping characteristics of the mechanism were examined with high-speed video II. EXPERIMENTAL METHOD camera system. A. Microscopic Observations Index Terms—Jumping Mechanism, Leaping Organ, Globular springtails are small insects that reach Springtail, Morphology, Jumping Characteristics around 1 mm in size. They have the jumping organ (furcula), which can be folded under the abdomen. Muscular action releasing the furcula can throw the I. INTRODUCTION insect well out of the way of predators. Jumping in insect movement is an effective way to In this paper, microscopic observations of the furcula escape predators, find food, and change locations. were conducted using the confocal laser scanning Grasshoppers, fleas, bush crickets, katydids, and locusts microscope. Confocal laser scanning microscopy is are particularly well known for jumping mechanisms to fluorescence – imaging technique that produces move around. -

Occasional Invaders
This publication is no longer circulated. It is preserved here for archival purposes. Current information is at https://extension.umd.edu/hgic HG 8 2000 Occasional Invaders Centipedes Centipede Millipede doors and screens, and by removal of decaying vegetation, House Centipede leaf litter, and mulch from around the foundations of homes. Vacuum up those that enter the home and dispose of the bag outdoors. If they become intolerable and chemical treatment becomes necessary, residual insecticides may be Centipedes are elongate, flattened animals with one pair of used sparingly. Poisons baits may be used outdoors with legs per body segment. The number of legs may vary from caution, particularly if there are children or pets in the home. 10 to over 100, depending on the species. They also have A residual insecticide spray applied across a door threshold long jointed antennae. The house centipede is about an inch may prevent the millipedes from entering the house. long, gray, with very long legs. It lives outdoors as well as indoors, and may be found in bathrooms, damp basements, Sowbugs and Pillbugs closets, etc. it feeds on insects and spiders. If you see a centipede indoors, and can’t live with it, escort it outdoors. Sowbugs and pillbugs are the only crustaceans that have Centipedes are beneficial by helping controlArchived insect pests and adapted to a life on land. They are oval in shape, convex spiders. above, and flat beneath. They are gray in color, and 1/2 to 3/4 of an inch long. Sowbugs have two small tail-like Millipedes appendages at the rear, and pillbugs do not. -

Springtails1 P
ENY-228 Springtails1 P. G. Koehler, M. L. Aparicio, and M. Pfiester2 organic material and other nutrients to return to the soil; these nutrients are later used by plants. Occasionally, springtails attack young seedlings and may damage the roots and stems. Biology and Description The Order Collembola has two suborders easily distin- guished by their body shape. One appears linear (Figure 1), while the other appears globular (Figure 2). These suborders are further divided into families that contain the separate species. Only seven families include the 650 species in North America. Worldwide, 3,600 species have been discovered. This fact sheet is included in SP134: Pests in and around the Florida Home, which is available from the UF/IFAS Extension Bookstore. http:// ifasbooks.ifas.ufl.edu/p-154-pests-in-andaroundthe-florida-home.aspx Figure 1. A linear springtail. Springtails are minute insects without wings in the Order Springtails range in length from 0.25 to 6 mm, but are Collembola. They occur in large numbers in moist soil and normally about 1 mm long. Colors range from white to can be found in homes with high humidity, organic debris, yellow, gray, or blue-gray. Attached to the tip of the abdo- or mold. Homeowners sometimes discover these insects men is a forked appendage resembling a lever and called occurring in large numbers in swimming pools, potted the furcula. At rest, a clasp called the tenaculum holds the plants, or in moist soil and mulch. They feed on fungi, furcula to the abdomen. When disturbed or threatened, fungal spores, and decaying, damp vegetation, causing the springtail’s tenaculum releases the furcula, which then 1. -

De Springstaarten Van Nederland: Het Genus ORCHESELLA (Hexapoda: Entognatha: Collembola) Matty Berg
de springstaarten van nederland: het genus ORCHESELLA (hexapoda: entognatha: collembola) Matty Berg Springstaarten kruipen al minstens 450 miljoen jaar op aarde rond en komen in bijna elk ecosysteem voor. Ze zijn vaak in grote aantallen aanwezig en hebben dan een zeer belangrijke functie. Enerzijds zijn ze betrokken bij de afbraak van organisch materiaal en anderzijds als voedsel voor allerlei predatoren. Uit Nederland zijn ruim 200 soorten springstaarten bekend en nog tientallen soorten zijn te verwachten. Een samenvatting van de kennis is zeer tijdrovend. Daarom is besloten tot een reeks artikelen, waarbij steeds een of meerdere genera worden behandeld, met een tabel tot de soorten, ver- spreidingskaarten en een ecologisch profiel. In dit eerste artikel wordt het genus Orchesella behandeld, met vier bekende soorten in ons land. inleiding nieuwe soorten gevonden voor onze fauna. De Al sinds 1887 wordt gepubliceerd over het voorko- nieuwe soorten worden genoemd in de hand- men van springstaarten in ons land. In dat jaar leiding voor het karteren van springstaarten publiceerde Oudemans in zijn proefschrift een (Berg 2002), maar de vondsten worden hier naamlijst van de Nederlandse Collembola, met niet onderbouwd met bewijsmateriaal en nadere 36 soorten. Pas in 1930 verschijnt een nieuwe gegevens. Het is dus hoog tijd voor een kritische lijst met 54 soorten en 17 variëteiten (Buitendijk naamlijst van de Nederlandse springstaarten. 1930). Een aanvulling komt elf jaar later (Buiten- dijk 1941), met 62 soorten en 16 variëteiten. Dit Een volledige revisie van de Nederlandse naam- is de laatste officiële naamlijst van springstaarten lijst is zeer tijdrovend, zeker als elke nieuwe die voor ons land is verschenen. -

Data Providers
Using Biodiversity Data from the NBN Database for Research y Paula Lightfoot, NBN Trust Data Access Officer Introduction to the NBN Database 1. Overview of available data 2. Finding and accessing data 3. Evaluating data quality 4. Using and referencing data http://data.nbn.org.uk Summary of Available Data • 91 million georeferenced taxon occurrence records. • 27 habitat datasets and 44 site boundary datasets to provide context and act as filters. • 856 datasets from 150 data providers. • Standard data format. • Standard taxonomy from UK Species Inventory. http://data.nbn.org.uk Data Providers Records in the NBN Database by data provider type. January 2014 (n = 91,206,588) • A large proportion of data comes from skilled amateur naturalists. • Data collated taxonomically and/or geographically. • Some structured surveys, much ad hoc recording. Geographic Coverage and Sampling Effort: • Recorder effort and data mobilisation are not evenly spread across the British Isles. • New NBN Gateway (v.5) extends coverage to include the Channel Islands and offshore data. • National Biodiversity Data Centre is the repository for ROI data. Sampling Effort Collembola Recording Scheme BTO Second Atlas of Breeding Birds 10,633 records of 336 species in Britain and Ireland: 1988-1991 over 200 years 1,465,400 records of 272 species over 4 years Sampling Effort Orchesella villosa (a springtail) http://tombio.myspecies.info/ Taxonomic Coverage Taxonomic breakdown of records in the NBN Database at January 2014 n = 91,269,685 Currency of Data Number of records in the NBN Database by year of record (January 2014) n = 89,091,428 (98% of total) Data Attributes Standard attributes in NBN Exchange Format: Required: Unique record key, taxon, date, date type, coordinates/grid reference/polygon, projection, precision (what? where? when?) Optional: Survey key, sample key, absent, sensitive, site key, site name, recorder, determiner Other attributes are not (yet) standardised across datasets: e.g. -

Five New Species of Orchesella (Collembola: Entomobryidae)
Proceedings of the Iowa Academy of Science Volume 84 Number Article 3 1977 Five New Species of Orchesella (Collembola: Entomobryidae) K. A. Christiansen Grinnell College B. E. Tucker Grinnell College Let us know how access to this document benefits ouy Copyright ©1977 Iowa Academy of Science, Inc. Follow this and additional works at: https://scholarworks.uni.edu/pias Recommended Citation Christiansen, K. A. and Tucker, B. E. (1977) "Five New Species of Orchesella (Collembola: Entomobryidae)," Proceedings of the Iowa Academy of Science, 84(1), 1-13. Available at: https://scholarworks.uni.edu/pias/vol84/iss1/3 This Research is brought to you for free and open access by the Iowa Academy of Science at UNI ScholarWorks. It has been accepted for inclusion in Proceedings of the Iowa Academy of Science by an authorized editor of UNI ScholarWorks. For more information, please contact [email protected]. Q Christiansen and Tucker: Five New Species of Orchesella (Collembola: Entomobryidae) \ \ (-:f-6 v,tff rlD' l Five New Species of Orchesella (Collembola: Entomobryidae) C ·if K. A. CHR1STIANSEN 1 and B. E. TUCKER2 CHRISTIANSEN, K. A. and BRUCE E. TUCKER (Dept. of Biology, bryidae) new to science . The chaetotaxy of the abdomen as well as the antenna! Grinnell College, Grinnell IA 50112). Five New Species of Orchesella (Col pin seta are used systematically for the first time in the taxonomy of the genus. lembola: Entomobryidae). Proc. Iowa Acad. Sci. 84(1): 1-13, 1977 . INDEX DESCRIPTORS: Collembola Taxonomy, North American Insect This paper describes 5 species of the genus Orchese/la (Collembola: Entomo- Taxonomy. -
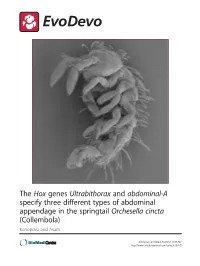
Abdominal-A Specify Three Different Types of Abdominal Appendage in the Springtail Orchesella Cincta (Collembola) Konopova and Akam
The Hox genes Ultrabithorax and abdominal-A specify three different types of abdominal appendage in the springtail Orchesella cincta (Collembola) Konopova and Akam Konopova and Akam EvoDevo 2014, 5:2 http://www.evodevojournal.com/content/5/1/2 Konopova and Akam EvoDevo 2014, 5:2 http://www.evodevojournal.com/content/5/1/2 RESEARCH Open Access The Hox genes Ultrabithorax and abdominal-A specify three different types of abdominal appendage in the springtail Orchesella cincta (Collembola) Barbora Konopova* and Michael Akam Abstract Background: In Drosophila and many other insects, the Hox genes Ultrabithorax (Ubx) and abdominal-A (abd-A) suppress limb formation on most or all segments of the abdomen. However, a number of basal hexapod lineages retain multiple appendages on the abdomen. In the collembolans or springtails, three abdominal segments develop specialized organs that originate from paired appendage primordia which fuse at the midline: the first abdominal segment bears the collophore (ventral tube), involved in osmoregulation; the fourth segment bears the furca, the leaping organ, and the third segment bears the retinaculum, which retains the furca at rest. Ubx and abd-A are known to be expressed in the springtail abdomen, but what role they play in specifying these distinct abdominal appendages is not known. This is largely because no genetic model has been established in collembolans or any other non-insect hexapod. Results: We have developed a convenient method for laboratory culture of the collembolan Orchesella cincta on defined media, a method for in-situ hybridization to embryos and a procedure for gene knockdown by parental injection of double-stranded RNA (RNAi). -

New Records of Springtail Fauna (Hexapoda: Collembola: Entomobryomorpha) from Ordu Province in Turkey
Turkish Journal of Zoology Turk J Zool (2017) 41: 24-32 http://journals.tubitak.gov.tr/zoology/ © TÜBİTAK Research Article doi:10.3906/zoo-1509-28 New records of springtail fauna (Hexapoda: Collembola: Entomobryomorpha) from Ordu Province in Turkey 1 2, 3 Muhammet Ali ÖZATA , Hasan SEVGİLİ *, Igor J. KAPRUS 1 Demir Karamancı Anatolian High School, Melikgazi, Kayseri, Turkey 2 Department of Biology, Faculty of Arts and Sciences, Ordu University, Ordu, Turkey 3 State Museum of Natural History, Ukrainian National Academy of Sciences, L’viv, Ukraine Received: 14.09.2015 Accepted/Published Online: 27.04.2016 Final Version: 25.01.2017 Abstract: This study aims to elucidate the Collembola fauna of the province of Ordu, which is situated between the Middle and Eastern Black Sea regions of Turkey. Although a large number of Collembolan specimens had been collected, only Entomobryomorpha species were given emphasis. From 44 different sampled localities of the province of Ordu, we recorded 6 families, 14 genera, and 28 species. Six of these species were previously recorded and 20 of them are new records for Turkey. The results were not surprising, considering that the sampled region had not been studied previously, quite like many habitats in Turkey. With our 20 new records (Entomobryomorpha), the grand total of the springtail fauna of Turkey is increased to 73 species. This represents an increase of almost 40% of the current list of known species. These numbers show us that the diversity of Collembola in Turkey is not thoroughly known and it is clear that numerous species remain undiscovered or undescribed. -

<I>Orchesella</I> (Collembola: Entomobryomorpha
University of Tennessee, Knoxville TRACE: Tennessee Research and Creative Exchange Masters Theses Graduate School 5-2015 A Molecular and Morphological Investigation of the Springtail Genus Orchesella (Collembola: Entomobryomorpha: Entomobryidae) Catherine Louise Smith University of Tennessee - Knoxville, [email protected] Follow this and additional works at: https://trace.tennessee.edu/utk_gradthes Part of the Entomology Commons Recommended Citation Smith, Catherine Louise, "A Molecular and Morphological Investigation of the Springtail Genus Orchesella (Collembola: Entomobryomorpha: Entomobryidae). " Master's Thesis, University of Tennessee, 2015. https://trace.tennessee.edu/utk_gradthes/3410 This Thesis is brought to you for free and open access by the Graduate School at TRACE: Tennessee Research and Creative Exchange. It has been accepted for inclusion in Masters Theses by an authorized administrator of TRACE: Tennessee Research and Creative Exchange. For more information, please contact [email protected]. To the Graduate Council: I am submitting herewith a thesis written by Catherine Louise Smith entitled "A Molecular and Morphological Investigation of the Springtail Genus Orchesella (Collembola: Entomobryomorpha: Entomobryidae)." I have examined the final electronic copy of this thesis for form and content and recommend that it be accepted in partial fulfillment of the requirements for the degree of Master of Science, with a major in Entomology and Plant Pathology. John K. Moulton, Major Professor We have read this thesis and recommend its acceptance: Ernest C. Bernard, Juan Luis Jurat-Fuentes Accepted for the Council: Carolyn R. Hodges Vice Provost and Dean of the Graduate School (Original signatures are on file with official studentecor r ds.) A Molecular and Morphological Investigation of the Springtail Genus Orchesella (Collembola: Entomobryomorpha: Entomobryidae) A Thesis Presented for the Master of Science Degree The University of Tennessee, Knoxville Catherine Louise Smith May 2015 Copyright © 2014 by Catherine Louise Smith All rights reserved.