Fourth International Circumpolar Arctic Vegetation Mapping Workshop
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Willows of Interior Alaska
1 Willows of Interior Alaska Dominique M. Collet US Fish and Wildlife Service 2004 2 Willows of Interior Alaska Acknowledgements The development of this willow guide has been made possible thanks to funding from the U.S. Fish and Wildlife Service- Yukon Flats National Wildlife Refuge - order 70181-12-M692. Funding for printing was made available through a collaborative partnership of Natural Resources, U.S. Army Alaska, Department of Defense; Pacific North- west Research Station, U.S. Forest Service, Department of Agriculture; National Park Service, and Fairbanks Fish and Wildlife Field Office, U.S. Fish and Wildlife Service, Department of the Interior; and Bonanza Creek Long Term Ecological Research Program, University of Alaska Fairbanks. The data for the distribution maps were provided by George Argus, Al Batten, Garry Davies, Rob deVelice, and Carolyn Parker. Carol Griswold, George Argus, Les Viereck and Delia Person provided much improvement to the manuscript by their careful editing and suggestions. I want to thank Delia Person, of the Yukon Flats National Wildlife Refuge, for initiating and following through with the development and printing of this guide. Most of all, I am especially grateful to Pamela Houston whose support made the writing of this guide possible. Any errors or omissions are solely the responsibility of the author. Disclaimer This publication is designed to provide accurate information on willows from interior Alaska. If expert knowledge is required, services of an experienced botanist should be sought. Contents -

"National List of Vascular Plant Species That Occur in Wetlands: 1996 National Summary."
Intro 1996 National List of Vascular Plant Species That Occur in Wetlands The Fish and Wildlife Service has prepared a National List of Vascular Plant Species That Occur in Wetlands: 1996 National Summary (1996 National List). The 1996 National List is a draft revision of the National List of Plant Species That Occur in Wetlands: 1988 National Summary (Reed 1988) (1988 National List). The 1996 National List is provided to encourage additional public review and comments on the draft regional wetland indicator assignments. The 1996 National List reflects a significant amount of new information that has become available since 1988 on the wetland affinity of vascular plants. This new information has resulted from the extensive use of the 1988 National List in the field by individuals involved in wetland and other resource inventories, wetland identification and delineation, and wetland research. Interim Regional Interagency Review Panel (Regional Panel) changes in indicator status as well as additions and deletions to the 1988 National List were documented in Regional supplements. The National List was originally developed as an appendix to the Classification of Wetlands and Deepwater Habitats of the United States (Cowardin et al.1979) to aid in the consistent application of this classification system for wetlands in the field.. The 1996 National List also was developed to aid in determining the presence of hydrophytic vegetation in the Clean Water Act Section 404 wetland regulatory program and in the implementation of the swampbuster provisions of the Food Security Act. While not required by law or regulation, the Fish and Wildlife Service is making the 1996 National List available for review and comment. -

Nornickel and the Kola Peninsula
THE BELLONA FOUNDATION Nornickel and the Kola Peninsula Photo: Thomas Nilsen ENVIRONMENTAL RESPONSIBILITY IN THE YEAR OF ECOLOGY JANUARY 2018 The Bellona Foundation is an international environmental NGO based in Norway. Founded in 1986 as a direct action protest group, Bellona has become a recognized technology and solution- oriented organizations with offices in Oslo, Brussels, Kiev, St. Petersburg and Murmansk. Altogether, some 60 engineers, ecologists, nuclear physicists, economists, lawyers, political scientists and journalists work at Bellona. Environmental change is an enormous challenge. It can only be solved if politicians and legislators develop clear policy frameworks and regulations for industry and consumers. Industry plays a role by developing and commercializing environmentally sound technology. Bellona strives to be a bridge builder between industry and policy makers, working closely with the former to help them respond to environmental challenges in their field, and proposing policy measures that promote new technologies with the least impact on the environment. Authors: Oskar Njaa © Bellona 201 8 Design: Bellona Disclaimer: Bellona endeavors to ensure that the information disclosed in this report is correct and free from copyrights, but does not warrant or assume any legal liability or responsibility for the accuracy, completeness, interpretation or usefulness of the information which may result from the use of this report. Contact: [email protected] Web page: www.bellona.org 1 Table of Contents 1 Introduction: ...................................................................................................................... -

Atlas of Rare Endemic Vascular Plants of the Arctic
Atlas of Rare Endemic Vascular Plants of the Arctic Technical Report No. 3 About CAFF Theprogram for the Conservation of Arctic Flora and Fauna (CAFF) of the Arctic Council was established lo address the special needs of Arctic ecosystems, species and thcir habitats in the rapid ly developing Arctic region. Itwas initiated as one of'four programs of the Arctic Environmental Protcction Strategy (AEPS) which was adopted by Canada, Denmark/Greenland, Finland, lceland, Norway, Russia, Swcdcn and the United States through a Ministeria! Declaration at Rovaniemi, Finland in 1991. Other programs initi ated under the AEPS and overlaken hy the Are.tie Council are the ArcticMonitoring and assessment Programme (AMAP), the program for Emergency Prevention, Preparcd ness and Response (EPPR) and the program for Protection of the Arctic Marine Envi ronment (PAME). Sinceits inaugural mccti.ng in Ottawa, Canada in 1992, the CAFF program has provided scientists, conscrvation managers and groups, and indigenous people of the north with a distinct forum in which lo tackle a wide range of Arctic conservation issues at the cir cumpolar level. CAFF's main goals, which are achieved in keeping with the concepts of sustainable developrnertt and utilisation, are: • to conserve Arctic Jlora and fauna, thcir diversity and thcir habitats; • to protect the Arctic ecosystems from threats; • to improve conservation management laws, reg ulations and practices for the Arclic; • to integrale Arctic interests into global conservation fora. CAFF operates rhrough a system of Designated Agencies and National Representatives responsible for CAFF in thcir rcspcctivc countries. CAFF also has an International Work ing Group wh.ith has met annually to assess progrcss and to develop Annual WorkPlans. -

FOH Newsletter 2012
FRIENDS Of The University Of Montana HERBARIUM Spring 2012 ....Bruce McCune MORE THAN VASCULAR AT MONTU By Andrea Pipp While perusing the vascular plant, moss, and lichen were the ecological relatives of mosses and incorporated specimens at MONTU you might see the name of Bruce them into his work. He relied on Mason Hale’s first edi- McCune. During his years at UM (1971-1979) Bruce con- tion of How to Know the Lichens and a long paper with tributed mounts of 62 vascular plant species and a larger keys titled “Lichens of the State of Washington” by Grace number of moss and lichen vouchers to the herbarium. Howard. He soon realized Hale’s book was very biased Bruce came to Missoula from the Midwest. Having toward eastern species, for even the most conspicuous grown up in Cincinnati and after completing his freshman lichens found on Mt. Sentinel weren’t in the book. Later year at Lawrence University in Appleton, Wisconsin, he discovered that Howard’s key contained as much mis- Bruce attended a wilderness botany class in northern Min- (Continued on page 7) nesota. There in the moss-carpeted forests he became quite interested in mosses and, in his words, was swept away by a remarkable young lady (Patricia Muir). Eager to escape the industrial winter of Appleton, Bruce fol- lowed Patricia to the University of Montana. That was 1971. The ‘new’ library was under construction, Hitch- cock and Cronquist, as a book, did not exist, and Botany stood on its own – a vibrant, active department, firmly ensconced in its own building. -

With Nansen in the North; a Record of the Fram Expedition in 1893-96
with-nXT^sen Ulllii I Hi nil I li "'III"!!!!!!!! Lieut JOHANSi; i*ll III Hi!; :1 III I iiil |i;iii'i'iiiiiiii i; \Ki THE LIBRARY OF THE UNIVERSITY OF CALIFORNIA LOS ANGELES WITH NANSEN IN THE NORTH [LANCASTER. K lOHANSEN. {Frontis/iiece. WITH NANSEN IN THE NORTH A Record of the Fram Expedition in 1893-96 BY HJALMAR JOHANSEN lieutenant in the norwegian army Translated from the Norwegian BY H. L. BR.EKSTAD LONDON WARD, LOCK AND CO LIMITED NEW YORK AND MELBOURNE 1899 — ijr Contents CHAPTER I TAGE The Equipment of the Expedition— Its Start—The Voyage along the Coast—Farewell to Norway I CHAI'TER II The First Ice — Arrival at Khabarova—Meeting with Trontheim Arrival of the Dogs— Life among the Samoyedes— Christofersen Leaves us—Excursion on Yalmal—The last Human Beings we saw lo CHAPTER III A Heavy Sea—Sverdrup Island—A Reindeer Hunt—The First Bear —A Stiff Pull—Firing with Kerosene 20 CHAPTER IV Death among the Dogs — Taimur Island — Cape Butterless — The Northernmost Point of the Old World—A Walrus Hunt—To the North 28 CHAPTER V Open Water—Unwelcome Guests—Fast in the Ice—Warping—The Northern Lights 34 CHAPTER VI First Day of Rest— Surprised by Bears—The Dogs are let Loose- Ice Pressure—A Hunt in the Dark 4° CHAPTER VII More Bears—The Power of Baking Powder— "Johansen's Friend"— V Electric Light— Shooting Competition ...•• 5° V 1212604 vi COXTENTS CHAPTER VIII PAGE Foot-races on the Ice—More about the Dogs—The Northern Lights- Adulterated Beer—Ice Pressure—Peder Attacked by a Bear . -

Sverdrup-Among-The-Tundra-People
AMONG THE TUNDRA PEOPLE by HARALD U. SVERDRUP TRANSLATED BY MOLLY SVERDRUP 1939 Copyright @ 1978 by Regents of the University of California. All rights reserved. No part of this book may be reproduced or utilized in any form or by any means, elec- tronic or mechanical, including photocopying, recording, or by any information storage and retrieval system, without permission in writing from the regents. Distributed by : Scripps Institution of Oceanography A-007 University of California, San Diego La Jolla, California 92093 Library of Congress # 78-60483 ISBN # 0-89626-004-6 ACKNOWLEDGMENTS We are indebted to Molly Sverdrup (Mrs. Leif J.) for this translation of Hos Tundra-Folket published by Gyldendal Norsk Forlag, Oslo, 1938. We are also indebted to the late Helen Raitt for recovering the manuscript from the archives of the Scripps Institution of Oceanography. The Norwegian Polar Institute loaned negatives from Sverdrup's travels among the Chukchi, for figures 1 through 4. Sverdrup's map of his route in the Chukchi country in 19 19/20 was copied from Hos Tundra-Folket. The map of the Chukchi National Okrug was prepared by Fred Crowe, based on the American Geographic Society's Map of the Arctic Region (1975). The map of Siberia was copied from Terence Armstrong's Russian Settlement in the North (1 965) with permission of the Cambridge University Press. Sam Hinton drew the picture of a reindeer on the cover. Martin W. Johnson identified individuals in some of the photographs. Marston C Sargent Elizabeth N. Shor Kittie C C Kuhns Editors The following individuals, most of whom were closely associated with Sverdrup, out of respect for him and wishing to assure preservation of this unusual account, met part of the cost of publication. -

Euphorbiaceae)
Polish Botanical Journal 60(2): 147–161, 2015 DOI: 10.1515/pbj-2015-0024 PHYTOGEOGRAPHICAL ANALYSIS OF EUPHORBIA SUBGENUS ESULA (EUPHORBIACEAE) Dmitry V. Geltman Abstract. Euphorbia subg. Esula is one of four major clades within the genus. A geographical analysis of the 466 species in the subgenus is reported here. Every species was assigned to one of 29 geographical elements clustered in ten groups of ele- ments. This geographical analysis showed that the Tethyan group (comprising nine geographical elements) clearly dominates the subgenus and contains 260 species (55.79% of the total number of species). The most numerous geographical elements are Irano-Turanian (105 species) and Mediterranean (85). Other significant groups of elements are Boreal (91 species, 19.54%), East Asian (40 species, 8.58%), Madrean (26 species, 5.58%), Paleotropical (23 species, 4.94%) and South African (16 species, 3.43%). The area of the Tethyan floristic subkingdom is the center of the modern diversity of E. subg. Esula. It is likely that such diversity is the result of intensive speciation that took place during the Eocene–Miocene. Key words: Euphorbia subg. Esula, geographical elements, Irano-Turanian floristic region, Mediterranean floristic region, phytogeographical analysis, Tethyan floristic subkingdom Dmitry V. Geltman, Komarov Botanical Institute of the Russian Academy of Sciences, Prof. Popov Street, 2, St. Petersburg, 197376, Russia; e-mail: [email protected] Introduction genus euphorbia and its taxonomy cantly differ from traditional ones. For subgenus Esula (Riina et al. 2013), 21 sections were ac- The giant genus Euphorbia L. (Euphorbiaceae) re- cepted on the basis of analyses of the combined cently became a subject of detailed phylogenetic and ITS + ndhF dataset (Fig. -
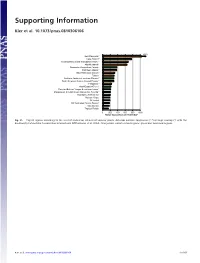
Supporting Information
Supporting Information Kier et al. 10.1073/pnas.0810306106 1350 New Caledonia** Cape Region** Polynesia-Micronesia and Eastern Pacific** Atlantic Islands* Queensland tropical rain forests Caribbean Islands** East Melanesian Islands** Taiwan Northern Andes incl. northern Páramo** South American Atlantic Coastal Forests** Philippines** West-Ecuador/Choco* Peruvian/Bolivian Yungas & montane forests* Madagascar & Indian Ocean Islands incl. Socotra** Mountains of SW China** Western Ghats* Sri Lanka* SW Australian Floristic Region** New Guinea Tropical Florida 0 200 400 600 800 1000 Range equivalents per 10,000 km² Fig. S1. Top 20 regions according to the level of endemism richness of vascular plants. Asterisks indicate congruence (**) or large overlap (*) with the biodiversity hotspots by Conservation International (Mittermeier, et al. 2004). Orange bars indicate island regions, green bars mainland regions. Kier et al. www.pnas.org/cgi/content/short/0810306106 1of15 Fig. S2. Endemism richness (range equivalents per 10,000 km2) of terrestrial vertebrates at the ecoregion level of mainland (green) and island regions (orange). Boxes mark second and third quartiles, whiskers the nonoutlier range of the data. (A) Terrestrial vertebrates, (B) amphibians, (C) birds, (D) reptiles, and (E) mammals. Kier et al. www.pnas.org/cgi/content/short/0810306106 2of15 Spearman rank correlation Vascular 0.75 0.82 0.73 0.78 0.83 plants 90 Amphibians 0.76 0.82 0.82 0.87 0 90 Reptiles 0.85 0.85 0.94 ER rank ER rank 0 90 Birds 0.89 0.96 0 90 Mammals 0.94 0 90 Tetrapods ER rank ER rank ER rank 0 090090090090090 ER rank ER rank ER rank ER rank ER rank Fig. -

Arctic National Wildlife Refuge Volume 2
Appendix F Species List Appendix F: Species List F. Species List F.1 Lists The following list and three tables denote the bird, mammal, fish, and plant species known to occur in Arctic National Wildlife Refuge (Arctic Refuge, Refuge). F.1.1 Birds of Arctic Refuge A total of 201 bird species have been recorded on Arctic Refuge. This list describes their status and abundance. Many birds migrate outside of the Refuge in the winter, so unless otherwise noted, the information is for spring, summer, or fall. Bird names and taxonomic classification follow American Ornithologists' Union (1998). F.1.1.1 Definitions of classifications used Regions of the Refuge . Coastal Plain – The area between the coast and the Brooks Range. This area is sometimes split into coastal areas (lagoons, barrier islands, and Beaufort Sea) and inland areas (uplands near the foothills of the Brooks Range). Brooks Range – The mountains, valleys, and foothills north and south of the Continental Divide. South Side – The foothills, taiga, and boreal forest south of the Brooks Range. Status . Permanent Resident – Present throughout the year and breeds in the area. Summer Resident – Only present from May to September. Migrant – Travels through on the way to wintering or breeding areas. Breeder – Documented as a breeding species. Visitor – Present as a non-breeding species. * – Not documented. Abundance . Abundant – Very numerous in suitable habitats. Common – Very likely to be seen or heard in suitable habitats. Fairly Common – Numerous but not always present in suitable habitats. Uncommon – Occurs regularly but not always observed because of lower abundance or secretive behaviors. -

Kola Oleneostrovskiy Grave Field: a Unique Burial Site in the European Arctic
Kola Oleneostrovskiy Grave Field: A Unique Burial Site in the European Arctic Anton I. Murashkin, Evgeniy M. Kolpakov, Vladimir Ya. Shumkin, Valeriy I. Khartanovich & Vyacheslav G. Moiseyev Anton I. Murashkin, Department of Archaeology, St Petersburg State University, Mendeleyevskaya liniya 5, RU-199034 St Petersburg, Russia: [email protected], [email protected] Evgeniy M. Kolpakov, Department of Palaeolithic, Institute for the History of Material Culture, Russian Academy of Sciences, Dvortsovaya nab. 18, RU-191186 St Petersburg, Russia: [email protected] Vladimir ya. Shumkin, Department of Palaeolithic, Institute for the History of Material Culture, Russian Academy of Sciences, Dvortsovaya nab. 18, RU-191186 St Petersburg, Russia: [email protected] Valeriy I. Khartanovich, Department of Anthropology, Peter the Great Museum of Anthropology and Ethnography (Kunstkamera), Russian Academy of Sciences, Universitetskaya nab. 3, RU-199034 St Petersburg, Russia: [email protected] Vyacheslav G. Moiseyev, Department of Anthropology, Peter the Great Museum of Anthropology and Ethnography (Kunstkamera), Russian Academy of Sciences, Universitetskaya nab. 3, RU-199034 St Petersburg, Russia: [email protected] Abstract The Kola Oleneostrovskiy grave field (KOG) is the main source of information for the physical and cultural anthropology of the Early Metal Period population of the Kola Peninsula and the whole northern Fennoscandia.1 Excavations were conducted here in 1925, 1928, 1947–1948, and 2001–2004 by A. Shmidt, N. Gurina, and V. Shumkin. Altogether 32 burials containing the remains of 43 individuals were investigated. During the excavations, also remains of wooden grave constructions were found. The site is exceptionally rich in burial goods, including numerous bone, antler, stone, ceramic, and bronze items. -

Introduction to Picor-Ice SNAME Presentation Notes Slide 1 – Title Thank You, SNAME Arctic, for Your Kind Invitation to Speak
Introduction to PicoR-Ice SNAME Presentation Notes Slide 1 – Title Thank you, SNAME Arctic, for your kind invitation to speak. This goes back to an excellent presentation earlier in the year (2018) by Bruce Calderbank on ice- related marine casualties in Canada. I asked if a blatant commercial presentation might be in order. Following last month’s update on the Arktos evacuation vehicle, the chairman invited me to deliver today’s presentation on PicoR-Ice. Thank you again. Slide 2 – The PicoR-Ice System PicoR-Ice is a ground-penetrating radar (GPR), the same technology you see on cable TV documentaries of treasure hunts and archaeological digs. But PicoR- Ice focuses on ice and snow thickness measurements. It is “non-invasive,” reducing need for drilling in ice. It processes radar returns and displays the underfoot reflection pattern instantly. And the entire system fits in a very manageable carrying bag, seen here on my back deck table with a standard champagne bottle for scale. Slide 3 – System Spec Sheet We have an engineering audience here today and so the system specifications are essential. A few highlights. Optimum ice thickness measurement down to 2 metres underfoot; snow layer thickness to 3 metres. Accurate to 2-3 cm. Transmission frequency of 1700 MHz trades off depth of penetration for increased resolution, important for operational underfoot thickness calculations. 30 to 60 pulses per second. When running vehicle-based survey, maximum vehicle speed of 40 km/h. The sensing technology is enclosed in a rugged and compact transmit-receive package (show actual module to audience).