Euphorbiaceae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

HAPARANDA MINISTERIAL DECLARATION on CIRCUMBOREAL COOPERATION on FORESTS Adopted at Haparanda, Sweden on June 26Th, 2018
HAPARANDA MINISTERIAL DECLARATION ON CIRCUMBOREAL COOPERATION ON FORESTS Adopted at Haparanda, Sweden on June 26th, 2018 We, the Ministers responsible for forests and their high-level representatives, from the circumboreal countries of Canada, Finland, Norway, Sweden, the Russian Federation and the United States of America, Noting that boreal forests grow in high-latitude environments in our respective countries, and that the boreal domain represents about 30% of the global forest area; Highlighting that boreal forests are important sources of renewable goods and services, and serve as habitat for a unique and vulnerable collection of biodiversity; Acknowledging that, in addition to local communities relying on the forest sector, boreal forests are home to many indigenous peoples who possess traditional knowledge and rely on forests to provide resources for their livelihoods, income, and cultural and spiritual values; Reiterating that countries in the circumboreal region are committed to sustainable forest management and report voluntarily on the environmental, social and economic status of their forests according to the criteria and indicators for sustainable forest management under the Montréal Process or Forest Europe; Recognizing that sustainable forest management in the circumboreal region can play a critical role in achieving the 2030 Agenda for Sustainable Development and its Sustainable Development Goals, as well as contribute to the implementation of other international outcomes including, as applicable, the Paris Agreement, -

Campanulaceae) Based on ITS and Tranl-F Sequence Data: Implications for a Reclassification
CORE Metadata, citation and similar papers at core.ac.uk Provided by University of the Western Cape Research Repository Cupido, C. N. et al. (2013). Phylogeny of Southern African and Australasian Wahlenbergioids (Campanulaceae) based on ITS and tranL-F sequence data: implications for a reclassification. Systematic Botany, 38(2): 523 – 535 http:// doi.org/10.1600/036364413X666714 dx. Phylogeny of Southern African and Australasian Wahlenbergioids (Campanulaceae) based on ITS and trnL-F sequence data: implications for a reclassification Christopher N. Cupido , Jessica M. Prebble , and William M. M. Eddie Abstract The Campanulaceae: Wahlenbergioideae currently comprises 15 genera, one of which, Wahlenbergia, is widespread over the southern continents. Southern Africa is the region with maximum wahlenbergioid diversity with 12 genera and approximately 252 species. A second center is Australasia with 38 Wahlenbergia species. This study used a broad sample of wahlenbergioid diversity from South Africa, Australia, and New Zealand to reconstruct a phylogeny based on chloroplast trnL-F and nuclear ITS sequences. Data were analyzed separately and in combination using parsimony and Bayesian methods. The results suggest that for the wahlenbergioids to be monophyletic Wahlenbergia hederacea has to be excluded and that none of the South African, Australian or New Zealand lineages are strictly monophyletic. There are five species assemblages that are in some disagreement with current classification in the family. Wahlenbergia, Prismatocarpus and Roella are shown to be non-monophyletic and implications for a reclassification are presented. Careful consideration of morphological characters is suggested before the adjustment of generic circumscriptions can be accomplished. Recent family-wide molecular phylogenetic studies have supported the view that the Campanulaceae s.s. -

The Plant Press the ARIZONA NATIVE PLANT SOCIETY
The Plant Press THE ARIZONA NATIVE PLANT SOCIETY Volume 36, Number 1 Summer 2013 In this Issue: Plants of the Madrean Archipelago 1-4 Floras in the Madrean Archipelago Conference 5-8 Abstracts of Botanical Papers Presented in the Madrean Archipelago Conference Southwest Coralbean (Erythrina flabelliformis). Plus 11-19 Conservation Priority Floras in the Madrean Archipelago Setting for Arizona G1 Conference and G2 Plant Species: A Regional Assessment by Thomas R. Van Devender1. Photos courtesy the author. & Our Regular Features Today the term ‘bioblitz’ is popular, meaning an intensive effort in a short period to document the diversity of animals and plants in an area. The first bioblitz in the southwestern 2 President’s Note United States was the 1848-1855 survey of the new boundary between the United States and Mexico after the Treaty of Guadalupe Hidalgo of 1848 ended the Mexican-American War. 8 Who’s Who at AZNPS The border between El Paso, Texas and the Colorado River in Arizona was surveyed in 1855- 9 & 17 Book Reviews 1856, following the Gadsden Purchase in 1853. Besides surveying and marking the border with monuments, these were expeditions that made extensive animal and plant collections, 10 Spotlight on a Native often by U.S. Army physicians. Botanists John M. Bigelow (Charphochaete bigelovii), Charles Plant C. Parry (Agave parryi), Arthur C. V. Schott (Stephanomeria schotti), Edmund K. Smith (Rhamnus smithii), George Thurber (Stenocereus thurberi), and Charles Wright (Cheilanthes wrightii) made the first systematic plant collection in the Arizona-Sonora borderlands. ©2013 Arizona Native Plant In 1892-94, Edgar A. Mearns collected 30,000 animal and plant specimens on the second Society. -

Euphorbia Subg
ФЕДЕРАЛЬНОЕ ГОСУДАРСТВЕННОЕ БЮДЖЕТНОЕ УЧРЕЖДЕНИЕ НАУКИ БОТАНИЧЕСКИЙ ИНСТИТУТ ИМ. В.Л. КОМАРОВА РОССИЙСКОЙ АКАДЕМИИ НАУК На правах рукописи Гельтман Дмитрий Викторович ПОДРОД ESULA РОДА EUPHORBIA (EUPHORBIACEAE): СИСТЕМА, ФИЛОГЕНИЯ, ГЕОГРАФИЧЕСКИЙ АНАЛИЗ 03.02.01 — ботаника ДИССЕРТАЦИЯ на соискание ученой степени доктора биологических наук САНКТ-ПЕТЕРБУРГ 2015 2 Оглавление Введение ......................................................................................................................................... 3 Глава 1. Род Euphorbia и основные проблемы его систематики ......................................... 9 1.1. Общая характеристика и систематическое положение .......................................... 9 1.2. Краткая история таксономического изучения и формирования системы рода ... 10 1.3. Основные проблемы систематики рода Euphorbia и его подрода Esula на рубеже XX–XXI вв. и пути их решения ..................................................................................... 15 Глава 2. Материал и методы исследования ........................................................................... 17 Глава 3. Построение системы подрода Esula рода Euphorbia на основе молекулярно- филогенетического подхода ...................................................................................................... 24 3.1. Краткая история молекулярно-филогенетического изучения рода Euphorbia и его подрода Esula ......................................................................................................... 24 3.2. Результаты молекулярно-филогенетического -

Bio 308-Course Guide
COURSE GUIDE BIO 308 BIOGEOGRAPHY Course Team Dr. Kelechi L. Njoku (Course Developer/Writer) Professor A. Adebanjo (Programme Leader)- NOUN Abiodun E. Adams (Course Coordinator)-NOUN NATIONAL OPEN UNIVERSITY OF NIGERIA BIO 308 COURSE GUIDE National Open University of Nigeria Headquarters 14/16 Ahmadu Bello Way Victoria Island Lagos Abuja Office No. 5 Dar es Salaam Street Off Aminu Kano Crescent Wuse II, Abuja e-mail: [email protected] URL: www.nou.edu.ng Published by National Open University of Nigeria Printed 2013 ISBN: 978-058-434-X All Rights Reserved Printed by: ii BIO 308 COURSE GUIDE CONTENTS PAGE Introduction ……………………………………......................... iv What you will Learn from this Course …………………............ iv Course Aims ……………………………………………............ iv Course Objectives …………………………………………....... iv Working through this Course …………………………….......... v Course Materials ………………………………………….......... v Study Units ………………………………………………......... v Textbooks and References ………………………………........... vi Assessment ……………………………………………….......... vi End of Course Examination and Grading..................................... vi Course Marking Scheme................................................................ vii Presentation Schedule.................................................................... vii Tutor-Marked Assignment ……………………………….......... vii Tutors and Tutorials....................................................................... viii iii BIO 308 COURSE GUIDE INTRODUCTION BIO 308: Biogeography is a one-semester, 2 credit- hour course in Biology. It is a 300 level, second semester undergraduate course offered to students admitted in the School of Science and Technology, School of Education who are offering Biology or related programmes. The course guide tells you briefly what the course is all about, what course materials you will be using and how you can work your way through these materials. It gives you some guidance on your Tutor- Marked Assignments. There are Self-Assessment Exercises within the body of a unit and/or at the end of each unit. -

Terrestrial Arthropods of Macaronesia
TERRESTRIAL ARTHROPODS OF MACARONESIA Biodiversity, Ecology and Evolution Title Terrestrial Artrhropods of Macaronesia - Biodiversity, Ecology and Evolution 1st edition, 2010 Editors Artur R. M. Serrano, Paulo A. V. Borges, Mário Boieiro and Pedro Oromí Sociedade Portuguesa de Entomologia Finantial support provided by Fundação para a Ciência e a Tecnologia, Portugal Project PDCT/BIA-BDE/59202/2004 Cover design by Security Print Cover photographs (and author): Desertas Islands (Photo by SPNM), Alloxantha fulva (Photo by P. Oromí) Misumena spinifera (Photo by P. Oromí), Guanchia uxoris (Photo by P. Oromí), Acrostira euphorbiae (Photo by P. Oromí), Dolichoiulus xylomistax (Photo by P. Oromí), Longitarsus isoplexidis (Photo by A. Serrano), Backcover photographs (and author): Selvagem Grande - Selvagens (Photo by SPNM), Turinyphia cavernicola (Photo by P. Borges), Herpisticus eremita (Photo by P. Oromí), Pseudoyersinia pilipes (Photo by P. Oromí), Hogna schmitzi (Photo by P. Oromí), Ischnura hastata (Photo by A. Cordero Ribera), Domene vulcanica (Photo by P. Oromí) Printed by Security Print - Sociedade de Indústria Gráfica, Lda. ISBN: 978-972-97241-2-1 Depósito Legal: XXX $POUFOUT __________________________________________________________________________________________ Preface by Antonio Machado INTRODUCTION Chapter 1 - The islands of Macaronesia, 1 J.M. Fernández-Palacios SECTION: BIODIVERSITY and CONSERVATION Chapter 2 - The provisional status of terrestrial arthropod inventories in the Macaronesian islands, 33 Jorge M. Lobo & Paulo A.V. Borges Chapter 3 - The Macaronesian province: patterns of species richness and endemism of arthropods, 49 Kostas A. Triantis, Paulo A.V. Borges, Joaquín Hortal & Robert J. Whittaker Chapter 4 - Patterns of Alpha and Beta Diversity of Epigean Arthropods at Contrasting Land-Uses of an Oceanic Island (Terceira, Azores), 73 Pedro Cardoso, Clara Gaspar, Francisco Dinis & Paulo A.V. -

The Lichen Flora of Gunib Plateau, Inner-Mountain Dagestan (North-East Caucasus, Russia)
Turkish Journal of Botany Turk J Bot (2013) 37: 753-768 http://journals.tubitak.gov.tr/botany/ © TÜBİTAK Research Article doi:10.3906/bot-1205-4 The lichen flora of Gunib plateau, inner-mountain Dagestan (North-East Caucasus, Russia) 1, 2 Gennadii URBANAVICHUS * , Aziz ISMAILOV 1 Institute of North Industrial Ecology Problems, Kola Science Centre, Russian Academy of Sciences, Apatity, Murmansk Region, Russia 2 Mountain Botanical Garden, Dagestan Scientific Centre, Russian Academy of Sciences, Makhachkala, Republic of Dagestan, Russia Received: 02.05.2012 Accepted: 15.03.2013 Published Online: 02.07.2013 Printed: 02.08.2013 Abstract: As a result of lichenological exploration of the Gunib plateau in the Republic of Dagestan (North-East Caucasus, Russia), we report 402 species of lichenised, 37 lichenicolous, and 7 nonlichenised fungi representing 151 genera. Nineteen species are recorded for the first time for Russia: Abrothallus chrysanthus J.Steiner, Abrothallus microspermus Tul., Caloplaca albopruinosa (Arnold) H.Olivier, Candelariella plumbea Poelt & Vězda, Candelariella rhodax Poelt & Vězda, Cladonia firma (Nyl.) Nyl., Halospora deminuta (Arnold) Tomas. & Cif., Halospora discrepans (J.Lahm ex Arnold) Hafellner, Lichenostigma epipolina Nav.-Ros., Calat. & Hafellner, Milospium graphideorum (Nyl.) D.Hawksw., Mycomicrothelia atlantica D.Hawksw. & Coppins, Parabagliettoa cyanea (A.Massal.) Gueidan & Cl.Roux, Placynthium garovaglioi (A.Massal.) Malme, Polyblastia dermatodes A.Massal., Rusavskia digitata (S.Y.Kondr.) S.Y.Kondr. & Kärnefelt, Squamarina stella-petraea Poelt, Staurothele elenkinii Oxner, Toninia nordlandica Th.Fr., and Verrucaria endocarpoides Servít. In addition, 71 taxa are new records for the Caucasus and 15 are new to Asia. Key words: Lichens, lichenicolous fungi, biodiversity, Gunib plateau, limestone, Dagestan, Caucasus, Russia 1. -

Final Environmental and Social Assessment Report (ESAR) and Environmental Management Plan (EMP) for Rehabilitation of Part of Th
Environmental and Social Assessment Report for Rehabilitation of part of the state road R2343 (old mark R-502), section Delchevo – Golak PUBLIC ENTERPRISE FOR STATE ROADS Republic of Macedonia Final Environmental and Social Assessment Report (ESAR) and Environmental Management Plan (EMP) for Rehabilitation of part of the state road R2343 (old mark R-502), section Delchevo – Golak - Final draft before disclosure - April 2015 1 Environmental and Social Assessment Report for Rehabilitation of part of the state road R2343 (old mark R-502), section Delchevo – Golak Contents 1. Executive Summary and Conclusion ......................................................................................................... 3 2. Policy, Legal and Administrative Framework ............................................................................................ 3 3. Project Description ........................................................................................................................................ 6 3.1. Location of the project ......................................................................................................................... 6 4. Envionmental and Social Baseline data .................................................................................................. 14 4.1. Geological composition, hydrogeology and pedology .................................................................. 14 4.2. Climate and Meteorology ................................................................................................................. -

Poaceae) De México Species Richness, Classification and a Checklist of the Grasses (Poaceae) of Mexico
Artículo de investigación Riqueza de especies, clasificación y listado de las gramíneas (Poaceae) de México Species richness, classification and a checklist of the grasses (Poaceae) of Mexico J. Gabriel Sánchez-Ken Resumen: Antecedentes y Objetivos: En 2006 se dio a conocer el catálogo de las gramíneas de México, pero desde entonces tanto la clasificación como la apa- rición de especies nuevas cambiaron las cifras de esta familia en México y en el mundo. El objetivo principal de este estudio fue actualizar la lista de taxa en México, reuniendo el mayor número posible de bases de datos en línea. Otros objetivos incluyeron la comparación de la riqueza de especies de la familia Poaceae entre los países con mayor diversidad, así como analizar la riqueza de sus taxa a nivel estatal y por categorías taxonómicas. Métodos: Se consultaron bases de datos en línea, tanto nacionales como extranjeras, que incluían ejemplares de referencia de México, se descarga- ron y se curaron, además de una confirmación visual de la identidad de un gran porcentaje de ejemplares. Se extrajeron los nombres de las especies, estados donde fueron colectadas y los acrónimos de los herbarios. La base resultante se complementó solo con bibliografía a partir de 2006 y la no incluida en el catálogo que mostraba ejemplares de respaldo. Resultados clave: Se revisaron 227,973 registros que contabilizaron, en total, 1416 taxa, que incluyen 215 géneros, 1312 especies y 214 categorías infraespecíficas. Del total de especies, 1092 se consideran nativas y 224 introducidas (cultivadas y/o malezas). Las especies endémicas de México son 304 (27.83%) o 311 (28.10%) si se consideran categorías inferiores. -

Illustrated Flora of East Texas Illustrated Flora of East Texas
ILLUSTRATED FLORA OF EAST TEXAS ILLUSTRATED FLORA OF EAST TEXAS IS PUBLISHED WITH THE SUPPORT OF: MAJOR BENEFACTORS: DAVID GIBSON AND WILL CRENSHAW DISCOVERY FUND U.S. FISH AND WILDLIFE FOUNDATION (NATIONAL PARK SERVICE, USDA FOREST SERVICE) TEXAS PARKS AND WILDLIFE DEPARTMENT SCOTT AND STUART GENTLING BENEFACTORS: NEW DOROTHEA L. LEONHARDT FOUNDATION (ANDREA C. HARKINS) TEMPLE-INLAND FOUNDATION SUMMERLEE FOUNDATION AMON G. CARTER FOUNDATION ROBERT J. O’KENNON PEG & BEN KEITH DORA & GORDON SYLVESTER DAVID & SUE NIVENS NATIVE PLANT SOCIETY OF TEXAS DAVID & MARGARET BAMBERGER GORDON MAY & KAREN WILLIAMSON JACOB & TERESE HERSHEY FOUNDATION INSTITUTIONAL SUPPORT: AUSTIN COLLEGE BOTANICAL RESEARCH INSTITUTE OF TEXAS SID RICHARDSON CAREER DEVELOPMENT FUND OF AUSTIN COLLEGE II OTHER CONTRIBUTORS: ALLDREDGE, LINDA & JACK HOLLEMAN, W.B. PETRUS, ELAINE J. BATTERBAE, SUSAN ROBERTS HOLT, JEAN & DUNCAN PRITCHETT, MARY H. BECK, NELL HUBER, MARY MAUD PRICE, DIANE BECKELMAN, SARA HUDSON, JIM & YONIE PRUESS, WARREN W. BENDER, LYNNE HULTMARK, GORDON & SARAH ROACH, ELIZABETH M. & ALLEN BIBB, NATHAN & BETTIE HUSTON, MELIA ROEBUCK, RICK & VICKI BOSWORTH, TONY JACOBS, BONNIE & LOUIS ROGNLIE, GLORIA & ERIC BOTTONE, LAURA BURKS JAMES, ROI & DEANNA ROUSH, LUCY BROWN, LARRY E. JEFFORDS, RUSSELL M. ROWE, BRIAN BRUSER, III, MR. & MRS. HENRY JOHN, SUE & PHIL ROZELL, JIMMY BURT, HELEN W. JONES, MARY LOU SANDLIN, MIKE CAMPBELL, KATHERINE & CHARLES KAHLE, GAIL SANDLIN, MR. & MRS. WILLIAM CARR, WILLIAM R. KARGES, JOANN SATTERWHITE, BEN CLARY, KAREN KEITH, ELIZABETH & ERIC SCHOENFELD, CARL COCHRAN, JOYCE LANEY, ELEANOR W. SCHULTZE, BETTY DAHLBERG, WALTER G. LAUGHLIN, DR. JAMES E. SCHULZE, PETER & HELEN DALLAS CHAPTER-NPSOT LECHE, BEVERLY SENNHAUSER, KELLY S. DAMEWOOD, LOGAN & ELEANOR LEWIS, PATRICIA SERLING, STEVEN DAMUTH, STEVEN LIGGIO, JOE SHANNON, LEILA HOUSEMAN DAVIS, ELLEN D. -

GPS AZORES Project
GPS AZORES Project Title: Geopolitical framework of the Macaronesia region Project Coordinator: Helena Calado (Universidade dos Azores) Work Package Coordinator: Juan Luis Suarez de Vivero (Universidad de Sevilla) Authors: Elisabetta Menini, Firdaous Halim, Daniela Gabriel, Juan Luis Suarez de Vivero, Helena Calado, Fabiana Moniz, Mario Caña Varona. Submission date: 31 August 2018 Acknowledgements: Christine Ladiero for helping with statistics Citation: Menini E., Halim F., Gabriel, D., Suarez de Vivero, JL., Calado, H., Moniz, F., Caña Varona, M. 2018. Geopolitical framework of the Macaronesia region. GPS Azores project: Ponta Delgada. Page 1 This project was financed in 85% by FEDER and in 15% with regional funds through the Programa Operacional Açores 2020 (Operational Program Azores 2020), in scope of the project « GPSAZORES -ACORES-01-0145-FEDER-00002». Page 2 Contents ACRONYMS 4 1. Maritime scenario characterisation: Main political, jurisdictional and socio-economic features 5 1.1 Political geography of the region 5 1.1.1 The regional context: Macaronesia 5 1.1.2 Countries and territories 12 1.2 Maritime space 16 1.2.1 Macaronesia in the context of the United Nations Convention of the Law of the Sea 16 1.2.2 Maritime jurisdictions 19 1.2.3 Maritime borders 21 1.3 The socio-economic context: exploitation and uses of the maritime space 26 1.3.1 Demography 26 1.3.2 Economic development 27 1.3.3 Tourism 29 1.3.1 Fisheries and aquaculture 31 1.3.2 Regional maritime geo-economics 33 1.3.3 Other relevant maritime economic activities 33 -
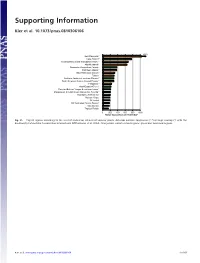
Supporting Information
Supporting Information Kier et al. 10.1073/pnas.0810306106 1350 New Caledonia** Cape Region** Polynesia-Micronesia and Eastern Pacific** Atlantic Islands* Queensland tropical rain forests Caribbean Islands** East Melanesian Islands** Taiwan Northern Andes incl. northern Páramo** South American Atlantic Coastal Forests** Philippines** West-Ecuador/Choco* Peruvian/Bolivian Yungas & montane forests* Madagascar & Indian Ocean Islands incl. Socotra** Mountains of SW China** Western Ghats* Sri Lanka* SW Australian Floristic Region** New Guinea Tropical Florida 0 200 400 600 800 1000 Range equivalents per 10,000 km² Fig. S1. Top 20 regions according to the level of endemism richness of vascular plants. Asterisks indicate congruence (**) or large overlap (*) with the biodiversity hotspots by Conservation International (Mittermeier, et al. 2004). Orange bars indicate island regions, green bars mainland regions. Kier et al. www.pnas.org/cgi/content/short/0810306106 1of15 Fig. S2. Endemism richness (range equivalents per 10,000 km2) of terrestrial vertebrates at the ecoregion level of mainland (green) and island regions (orange). Boxes mark second and third quartiles, whiskers the nonoutlier range of the data. (A) Terrestrial vertebrates, (B) amphibians, (C) birds, (D) reptiles, and (E) mammals. Kier et al. www.pnas.org/cgi/content/short/0810306106 2of15 Spearman rank correlation Vascular 0.75 0.82 0.73 0.78 0.83 plants 90 Amphibians 0.76 0.82 0.82 0.87 0 90 Reptiles 0.85 0.85 0.94 ER rank ER rank 0 90 Birds 0.89 0.96 0 90 Mammals 0.94 0 90 Tetrapods ER rank ER rank ER rank 0 090090090090090 ER rank ER rank ER rank ER rank ER rank Fig.