Effects of Nasal Obstruction on Maturation of the Jaw-Opening Reflex
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Comparison of Greater Palatine Nerve Block with Intravenous Fentanyl for Postoperative Analgesia Following Palatoplasty in Children
Jemds.com Original Research Article Comparison of Greater Palatine Nerve Block with Intravenous Fentanyl for Postoperative Analgesia Following Palatoplasty in Children Amol Singam1, Saranya Rallabhandi2, Tapan Dhumey3 1Department of Anaesthesiology, JNMC, DMIMS, Sawangi Meghe, Wardha Maharashtra, India. 2Department of Anaesthesiology, JNMC, DMIMS, Sawangi Meghe, Wardha, Maharashtra, India. 3Department of Anaesthesiology, JNMC, DMIMS, Sawangi Meghe, Wardha, Maharashtra, India. ABSTRACT BACKGROUND Good pain relief after palatoplasty is important as inadequate analgesia with vigorous Corresponding Author: cry leads to wound dehiscence, removal of sutures and extra nursing care. Decrease Dr. Saranya Rallabhandi, in oxygen requirement and cardio-respiratory demand occur with good pain relief Assisstant Professor, and also promotes early recovery. Preoperative opioids have concerns like sedation, Department of Anesthesiology, AVBRH, DMIMS (DU), Sawangi Meghe, respiratory depression and airway compromise. Greater palatine nerve block with Wardha- 442001, Maharashtra, India. bupivacaine is safe and effective without the risk of respiratory depression. The study E-mail: [email protected] was done to compare pain relief postoperatively with intravenous fentanyl and greater palatine nerve block in children following palatoplasty. DOI: 10.14260/jemds/2020/549 METHODS How to Cite This Article: 80 children of ASA I & II, between 1 to 7 years were included and allocated into two Singam A, Rallabhandi S, Dhumey T. Comparison of greater palatine nerve block groups of 40 each. Analgesic medication was given preoperatively after induction of with intravenous fentanyl for postoperative general anaesthesia, children in Group B received greater palatine nerve block with analgesia following palatoplasty in -1 2 mL 0.25% inj. Bupivacaine (1 mL on each side) and Group F received 2 μg Kg I.V. -
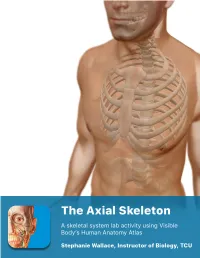
Lab Manual Axial Skeleton Atla
1 PRE-LAB EXERCISES When studying the skeletal system, the bones are often sorted into two broad categories: the axial skeleton and the appendicular skeleton. This lab focuses on the axial skeleton, which consists of the bones that form the axis of the body. The axial skeleton includes bones in the skull, vertebrae, and thoracic cage, as well as the auditory ossicles and hyoid bone. In addition to learning about all the bones of the axial skeleton, it is also important to identify some significant bone markings. Bone markings can have many shapes, including holes, round or sharp projections, and shallow or deep valleys, among others. These markings on the bones serve many purposes, including forming attachments to other bones or muscles and allowing passage of a blood vessel or nerve. It is helpful to understand the meanings of some of the more common bone marking terms. Before we get started, look up the definitions of these common bone marking terms: Canal: Condyle: Facet: Fissure: Foramen: (see Module 10.18 Foramina of Skull) Fossa: Margin: Process: Throughout this exercise, you will notice bold terms. This is meant to focus your attention on these important words. Make sure you pay attention to any bold words and know how to explain their definitions and/or where they are located. Use the following modules to guide your exploration of the axial skeleton. As you explore these bones in Visible Body’s app, also locate the bones and bone markings on any available charts, models, or specimens. You may also find it helpful to palpate bones on yourself or make drawings of the bones with the bone markings labeled. -

Morphometric Analysis of Greater Palatine Canal Via Cone-Beam Computed Tomography
DOI: 10.2478/bjdm-2018-0026 Y T E I C O S L BALKAN JOURNAL OF DENTAL MEDICINE A ISSN 2335-0245 IC G LO TO STOMA Morphometric Analysis of Greater Palatine Canal via Cone-Beam Computed Tomography SUMMARY Melih Özdede1, Elif Yıldızer Keriş2, Bülent Background/Aim: The morphology of the greater palatine canal (GPC) Altunkaynak3, İlkay Peker4 should be determined preoperatively to prevent possible complications in 1 Department of Dentomaxillofacial Radiology, surgical procedures required maxillary nerve block anesthesia and reduction Pamukkale University Faculty of Dentistry, of descending palatine artery bleeding. The purpose of this investigation was Denizli, Turkey to evaluate the GPC morphology. Material and Methods: In this retrospective 2 Canakkale Dental Hospital, Çanakkale, Turkey cross-sectional study, cone-beam computed tomography images obtained for 3 Department of Statistics, Gazi University various causes of 200 patients (females, 55%; males, 45%) age ranged between Faculty of Arts and Sciences, 18 and 86 (mean age±standard deviation=47±13.6) were examined. The mean Ankara, Turkey 4 Department of Dentomaxillofacial Radiology, length, mean angles of the GPC and anatomic routes of the GPC were evaluated. Gazi University Faculty of Dentistry, Results: The mean length of the GPC was found to be 31.07 mm and 32.01 mm Ankara, Turkey in sagittal and coronal sections, respectively. The mean angle of the GPC was measured as 156.16° and 169.23° in sagittal and coronal sections. The mean angle of the GPC with horizontal plane was measured as 113.76° in the sagittal sections and 92.94° in the coronal sections. -

Modeling the Jaw Mechanism of Pleuronichthys Verticalis: the Morphological Basis of Asymmetrical Jaw Movements in a Flatfish
JOURNAL OF MORPHOLOGY 256:1–12 (2003) Modeling the Jaw Mechanism of Pleuronichthys verticalis: The Morphological Basis of Asymmetrical Jaw Movements in a Flatfish Alice Coulter Gibb* Department of Ecology and Evolutionary Biology, University of California, Irvine, California ABSTRACT Several flatfish species exhibit the unusual water column for potential predators or prey. How- feature of bilateral asymmetry in prey capture kinemat- ever, the presence of both eyes on the same side of ics. One species, Pleuronichthys verticalis, produces lat- the head (i.e., the eyed side) also causes morpholog- eral flexion of the jaws during prey capture. This raises ical asymmetry of the skull and jaws (Yazdani, two questions: 1) How are asymmetrical movements gen- 1969). erated, and 2) How could this unusual jaw mechanism have evolved? In this study, specimens were dissected to Morphological asymmetry of the feeding appara- determine which cephalic structures might produce asym- tus creates the potential for another unusual verte- metrical jaw movements, hypotheses were formulated brate trait: asymmetry in jaw movements during about the specific function of these structures, physical prey capture. Two species of flatfish are known to models were built to test these hypotheses, and models exhibit asymmetrical jaw movements during prey were compared with prey capture kinematics to assess capture (Gibb, 1995, 1996), although the type of their accuracy. The results suggest that when the neuro- asymmetrical movement (i.e., kinematic asymme- cranium rotates dorsally the premaxillae slide off the try) is different in the two species examined. One smooth, rounded surface of the vomer (which is angled species, Xystreurys liolepis, produces limited kine- toward the blind, or eyeless, side) and are “launched” matic asymmetry during prey capture. -

Mandibular Jaw Movement and Masticatory Muscle Activity During Dynamic Trunk Exercise
dentistry journal Article Mandibular Jaw Movement and Masticatory Muscle Activity during Dynamic Trunk Exercise Daisuke Sugihara, Misao Kawara, Hiroshi Suzuki * , Takashi Asano, Akihiro Yasuda, Hiroki Takeuchi, Toshiyuki Nakayama, Toshikazu Kuroki and Osamu Komiyama Division of Oral Function and Rehabilitation, Department of Oral Health Science, Nihon University School of Dentistry at Matsudo, 870-1 Sakaecho, Nishi-2, Matsudo, Chiba 271-8587, Japan; [email protected] (D.S.); [email protected] (M.K.); [email protected] (T.A.); [email protected] (A.Y.); [email protected] (H.T.); [email protected] (T.N.); [email protected] (T.K.); [email protected] (O.K.) * Correspondence: [email protected]; Tel.: +81-47-360-9642 Received: 16 October 2020; Accepted: 30 November 2020; Published: 2 December 2020 Abstract: The examination of jaw movement during exercise is essential for an improved understanding of jaw function. Currently, there is no unified view of the mechanism by which the mandible is fixed during physical exercise. We hypothesized that during strong skeletal muscle force exertion in dynamic exercises, the mandible is displaced to a position other than the maximal intercuspal position and that mouth-opening and mouth-closing muscles simultaneously contract to fix the displaced mandible. Therefore, we simultaneously recorded mandibular jaw movements and masticatory muscle activities during dynamic trunk muscle force exertion (deadlift exercise) in 24 healthy adult males (age, 27.3 2.58 years). The deadlift was divided into three steps: ± Ready (reference), Pull, and Down. -

Pristipomoides Auricilla (Jordan, Evermann, and Tanaka, 1927) (Plate X, 67) Frequent Synonyms / Misidentifications: None / Other Species of Pristipomoides
click for previous page Perciformes: Percoidei: Lutjanidae 2909 Pristipomoides auricilla (Jordan, Evermann, and Tanaka, 1927) (Plate X, 67) Frequent synonyms / misidentifications: None / Other species of Pristipomoides. FAO names: En - Goldflag jobfish; Fr - Colas drapeau; Sp - Panchito abanderado. Diagnostic characters: Body elongate, laterally compressed. Nostrils on each side of snout close together. Jaws about equal or lower jaw protruding slightly. Premaxillae protrusible. Maxilla extending to vertical through anterior part of eye or slightly beyond. Upper and lower jaws both with an outer row of conical and canine teeth and an inner band of villiform teeth; vomer and palatines with teeth, those on vomer in triangular patch; no teeth on tongue. Maxilla without scales or longitudinal ridges. Interorbital region flattened. First gill arch with 8 to 11 gill rakers on upper limb, 17 to 21 on lower limb (total 27 to 32). Dorsal fin continuous, not deeply incised near junction of spinous and soft portions. Last soft ray of both dorsal and anal fins well produced, longer than next to last ray. Caudal fin forked. Pectoral fins long, equal to or somewhat shorter than head length. Dorsal fin with X spines and 11 soft rays. Anal fin with III spines and 8 soft rays. Pectoral-fin rays 15 or 16. Membranes of dorsal and anal fins without scales. Tubed lateral-line scales 67 to 74. Colour: body purplish or brownish violet; sides with numerous yellow spots or faint yellow chevron-shaped bands; dorsal fin yellowish to yellowish brown; upper lobe of caudal fin yellow. Sexual dichromatism: males over 27 cm (fork length) with much yellow on lower lobe of caudal fin, usually forming a distinct blotch; females with or without yellowish colour on lower lobe of caudal fin, but if yellow present, not forming a distinct blotch. -

Nasal Breathing No Tongue Habits Lip Seal Corruption of the “Big 3” Is the Root Cause of Nearly All Malocclusions
Timothy Gross, DMD Part One In the Physiologic Orthodontics class at the Las Vegas Institute, we focus heavily on the “Big 3.” Orthodontic treatment and post treatment orthodontic stability necessitate management and correction of the “Big 3.” So, what is the “Big 3?” 1 2 3 Nasal Breathing No Tongue Habits Lip Seal Corruption of the “Big 3” is the root cause of nearly all malocclusions. That’s right. Class I, Class II, and Class III malocclusions, crowding, deficient midfaces, cross bites, impacted teeth, high palatal arches, deep bites, open bites (and the list goes on) can all be explained by Enlow in his textbook, “The Essentials of Facial Growth.” That textbook is paramount for understanding normal facial growth and development and should be required reading for every dentist and dental specialist. Malocclusions, so predominant in our society, cannot be explained by genetics alone. Our phenotype, i.e. our genome in addition to environmental influences, is the result of corruption of the “Big 3.” The inability to breathe nasally, habitual mouth straight and make the upper and lower teeth couple breathing, and/or tongue habits lead to altered nicely. Unfortunately, the result is a skeletal and facial growth patterns which in turn lead to physiologic malocclusion with straight teeth, at observable dental and orthognathic malocclusions. the expense of the airway, the temporomandibular Orthodontic treatment is utilized to correct these joints and facial aesthetics. malocclusions, but the underlying pathophysiology must be addressed in order to achieve an optimal A Case Study… Me physiologic occlusion, balanced facial growth and As a 48 year old male, I had multiple issues: a facial beauty. -

A New Origin for the Maxillary Jaw
Developmental Biology 276 (2004) 207–224 www.elsevier.com/locate/ydbio A new origin for the maxillary jaw Sang-Hwy Leea, Olivier Be´dardb,1, Marcela Buchtova´b, Katherine Fub, Joy M. Richmanb,* aDepartment of Oral, Maxillofacial Surgery and Oral Science Research Center, Medical Science and Engineering Research Center, BK 21 Project for Medical Science, College of Dentistry Yonsei University, Seoul, Korea bDepartment of Oral Health Sciences, Faculty of Dentistry, University of British Columbia, Vancouver, BC, Canada, V6T 1Z3 Received for publication 7 April 2004, revised 5 August 2004, accepted 31 August 2004 Available online 5 October 2004 Abstract One conserved feature of craniofacial development is that the first pharyngeal arch has two components, the maxillary and mandibular, which then form the upper and lower jaws, respectively. However, until now, there have been no tests of whether the maxillary cells originate entirely within the first pharyngeal arch or whether they originate in a separate condensation, cranial to the first arch. We therefore constructed a fate map of the pharyngeal arches and environs with a series of dye injections into stage 13–17 chicken embryos. We found that from the earliest stage examined, the major contribution to the maxillary bud is from post-optic mesenchyme with a relatively minor contribution from the maxillo-mandibular cleft. Cells labeled within the first pharyngeal arch contributed exclusively to the mandibular prominence. Gene expression data showed that there were different molecular codes for the cranial and caudal maxillary prominence. Two of the genes examined, Rarb (retinoic acid receptor b) and Bmp4 (bone morphogenetic protein) were expressed in the post-optic mesenchyme and epithelium prior to formation of the maxillary prominence and then were restricted to the cranial half of the maxillary prominence. -

The Deep Structures of the Face
Thomas Jefferson University Jefferson Digital Commons Regional anatomy McClellan, George 1896 Vol. 1 Jefferson Medical Books and Notebooks November 2009 The Deep Structures of the Face Follow this and additional works at: https://jdc.jefferson.edu/regional_anatomy Part of the History of Science, Technology, and Medicine Commons Let us know how access to this document benefits ouy Recommended Citation "The Deep Structures of the Face" (2009). Regional anatomy McClellan, George 1896 Vol. 1. Paper 8. https://jdc.jefferson.edu/regional_anatomy/8 This Article is brought to you for free and open access by the Jefferson Digital Commons. The Jefferson Digital Commons is a service of Thomas Jefferson University's Center for Teaching and Learning (CTL). The Commons is a showcase for Jefferson books and journals, peer-reviewed scholarly publications, unique historical collections from the University archives, and teaching tools. The Jefferson Digital Commons allows researchers and interested readers anywhere in the world to learn about and keep up to date with Jefferson scholarship. This article has been accepted for inclusion in Regional anatomy McClellan, George 1896 Vol. 1 by an authorized administrator of the Jefferson Digital Commons. For more information, please contact: [email protected]. 136 THE DEEP STRUOTURES OF THE FAOE. in to the back part of the cavity; and the internal carotid ar tery and intern al jugular vein, with the hypoglossal, glosso-pharyngeal, and pneu mogastric nerves, were at the bottom of the wound, covered by a thin layer of fascia. THE DEEP STRUOTURES OF THE FAOE. The deep structures of the face, included in the pterygo-maxillary and superior maxillary regions, are of great surgical interest, owing to the importance of their relations and connections. -

Jaw Fracture Repair
Jaw Fractures What is a jaw fracture? This is when one of the bones of the upper or lower jaw breaks. Most commonly the lower jaw (the mandible) is broken, though the upper jaw (maxilla) and other bones of the face and orbit (eye socket) can be broken as well. Depending on the nature of the injury, multiple bones may be involved. Causes of jaw fractures can include trauma- such as fighting with another animal, being hit by a car or falling out of a window. Sometimes we see pathologic jaw fractures (due to disease rather than trauma) as a result of very severe periodontal disease or tumor (see our website for more information on what periodontal disease is, and how to treat it). Regardless of the cause, jaw fractures are painful, and require careful treatment planning in order to successfully treat the broken bone. The main goal of the Sacramento Veterinary Dentistry Services doctors is to give your pet the best possible quality of life. This means that we are always looking for the best, least invasive and most effective treatments. In terms of repairing a broken jaw, this would ideally mean using a non-invasive technique to repair the broken jaw that allows our patients to return to normal activities as soon as possible. There are many different repair techniques, and we will choose the best technique based on the dental x-ray images and overall oral condition of your pet. In most cases, this will involve wires and an intraoral splint made of plastic that binds to the teeth to immobilize the fracture and allow the bones to heal. -

Mandibulectomy/Resection of the Jaw
Mandibulectomy/Resection of the Jaw Mandibulectomy A mandibulectomy is a surgery to remove all or part of your jaw. Fibula free flap After part of your jaw is removed, your jaw may be rebuilt using bone from another part of your body, usually your leg. The bone is taken from your fibula, which is the smaller of the two bones in your lower leg. An artery, vein, and soft tissue will also be removed with the bone. This is called a fibula free flap. Skin graft You may also need a skin graft to cover your donor site. A skin graft is made by taking the top layer of skin from one part of your body and moving it to the surgical site that needs to be covered. If a skin graft is needed, it will be removed from another area, usually the buttock or thigh. Your surgeon will talk with you about the plan for your surgery. Who will be involved in your surgery? It is possible that multiple surgeons will be involved in your surgery. These surgical teams include the head and neck team, reconstruction team, and/or dental team. What happens during your surgery? Removal of the Disease During the surgery, the diseased bone will be removed from your jaw and the soft tissue around it. The bone and surrounding tissue will be sent to the Pathology Department for testing. This can take 3-4-hours. Once your head and neck surgeon has completed his or her part of the surgery, the reconstruction can be done. Reconstruction Your surgeon will transfer the tissue from your donor site (usually lower leg) to reconstruct your jaw. -

Glossary of Commonly Used Dental Terms
Glossary of Commonly Used Dental Terms A • Abutment: A tooth or implant used to support a prosthesis. A crown unit used as part of a fixed bridge. • Abscess: A localized inflammation due to a collection of pus in the bone or soft tissue, usually caused by an infection. • Amalgam: A dental filling material, composed of mercury and other minerals, used to fill decayed teeth. • Alveoloplasty: A surgical procedure used to recontour the supporting bone struc tures in preparation of a complete or partial denture. • Anesthetic: A class of drugs that eliminated of reduces pain. See local anesthetic. • Anterior: Refers to the teeth and tissues located towards the front of the mouth (upper or lower incisors and canines). • Apex: The tip end of a root. • Apexification: A method of inducing apical closure, or the continual apical develop ment of the root of an incompletely formed tooth, in which the pulp is no longer vital. B • Bicuspid: A two-cuspid tooth found between the molar and the cuspid also known as an eye tooth or canine tooth. • Biopsy: A process of removing tissue to determine the existence of pathology. • Bitewing x-ray: X-rays taken of the crowns of teeth to check for decay. • Bleaching: The technique of applying a chemical agent, usually hydrogen peroxide, to the teeth to whiten them. • Bondin: A process to chemically etch the tooth's enamel to better attach ( bond ) composite filling material, veneers, or plastic/acrylic. • Bone loss: The breakdown and loss of the bone that supports the teeth, usually caused by infection or long-term occlusal ( chewing areas of the teeth ) stress.