A Glucuronosyltransferase from Arabidopsis Thaliana Involved In
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Download (PDF)
Tomono et al.: Glycan evolution based on phylogenetic profiling 1 Supplementary Table S1. List of 173 enzymes that are composed of glycosyltransferases and functionally-linked glycan synthetic enzymes UniProt ID Protein Name Categories of Glycan Localization CAZy Class 1 Q8N5D6 Globoside -1,3-N -acetylgalactosaminyltransferase 1 Glycosphingolipid Golgi apparatus GT6 P16442 Histo-blood group ABO system transferase Glycosphingolipid Golgi apparatus GT6 P19526 Galactoside 2--L-fucosyltransferase 1 Glycosphingolipid Golgi apparatus GT11 Q10981 Galactoside 2--L-fucosyltransferase 2 Glycosphingolipid Golgi apparatus GT11 Q00973 -1,4 N -acetylgalactosaminyltransferase 1 Glycosphingolipid Golgi apparatus GT12 Q8NHY0 -1,4 N -acetylgalactosaminyltransferase 2 O -Glycan, N -Glycan, Glycosphingolipid Golgi apparatus GT12 Q09327 -1,4-mannosyl-glycoprotein 4--N -acetylglucosaminyltransferase N -Glycan Golgi apparatus GT17 Q09328 -1,6-mannosylglycoprotein 6--N -acetylglucosaminyltransferase A N -Glycan Golgi apparatus GT18 Q3V5L5 -1,6-mannosylglycoprotein 6--N -acetylglucosaminyltransferase B O -Glycan, N -Glycan Golgi apparatus GT18 Q92186 -2,8-sialyltransferase 8B (SIAT8-B) (ST8SiaII) (STX) N -Glycan Golgi apparatus GT29 O15466 -2,8-sialyltransferase 8E (SIAT8-E) (ST8SiaV) Glycosphingolipid Golgi apparatus GT29 P61647 -2,8-sialyltransferase 8F (SIAT8-F) (ST8SiaVI) O -Glycan Golgi apparatus GT29 Q9NSC7 -N -acetylgalactosaminide -2,6-sialyltransferase 1 (ST6GalNAcI) (SIAT7-A) O -Glycan Golgi apparatus GT29 Q9UJ37 -N -acetylgalactosaminide -2,6-sialyltransferase -

Flavonoid Glucodiversification with Engineered Sucrose-Active Enzymes Yannick Malbert
Flavonoid glucodiversification with engineered sucrose-active enzymes Yannick Malbert To cite this version: Yannick Malbert. Flavonoid glucodiversification with engineered sucrose-active enzymes. Biotechnol- ogy. INSA de Toulouse, 2014. English. NNT : 2014ISAT0038. tel-01219406 HAL Id: tel-01219406 https://tel.archives-ouvertes.fr/tel-01219406 Submitted on 22 Oct 2015 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Last name: MALBERT First name: Yannick Title: Flavonoid glucodiversification with engineered sucrose-active enzymes Speciality: Ecological, Veterinary, Agronomic Sciences and Bioengineering, Field: Enzymatic and microbial engineering. Year: 2014 Number of pages: 257 Flavonoid glycosides are natural plant secondary metabolites exhibiting many physicochemical and biological properties. Glycosylation usually improves flavonoid solubility but access to flavonoid glycosides is limited by their low production levels in plants. In this thesis work, the focus was placed on the development of new glucodiversification routes of natural flavonoids by taking advantage of protein engineering. Two biochemically and structurally characterized recombinant transglucosylases, the amylosucrase from Neisseria polysaccharea and the α-(1→2) branching sucrase, a truncated form of the dextransucrase from L. Mesenteroides NRRL B-1299, were selected to attempt glucosylation of different flavonoids, synthesize new α-glucoside derivatives with original patterns of glucosylation and hopefully improved their water-solubility. -

XAX1 from Glycosyltransferase Family 61 Mediates Xylosyltransfer to Rice Xylan
XAX1 from glycosyltransferase family 61 mediates xylosyltransfer to rice xylan Dawn Chiniquya,b, Vaishali Sharmab, Alex Schultinkc,d, Edward E. Baidoob,e, Carsten Rautengartenb, Kun Chengc,d, Andrew Carrollb, Peter Ulvskovf, Jesper Harholtf, Jay D. Keaslingb,e,g, Markus Paulyc,d, Henrik V. Schellerb,c,e, and Pamela C. Ronalda,b,h,1 aDepartment of Plant Pathology and the Genome Center, University of California, Davis, CA 95616; bJoint BioEnergy Institute, Emeryville, CA 94608; cDepartment of Plant and Microbial Biology, dEnergy Biosciences Institute, and gDepartment of Chemical and Biomolecular Engineering, Department of Bioengineering, University of California, Berkeley, CA 94720; ePhysical Biosciences Division, Lawrence Berkeley National Laboratory, Berkeley, CA 94720; fDepartment of Plant Biology and Biotechnology, University of Copenhagen, DK-1871 Frederiksberg C, Denmark; and hDepartment of Plant Molecular Systems Biotechnology and Crop Biotech Institute, Kyung Hee University, Yongin 446-701, Korea Edited by Diter von Wettstein, Washington State University, Pullman, WA, and approved August 31, 2012 (received for review February 6, 2012) Xylan is the second most abundant polysaccharide on Earth and Caulerpa that has β-1,3-D-xylan in place of cellulose, and the red represents an immense quantity of stored energy for biofuel pro- seaweeds Palmariales and Nemaliales that have a mixed linkage duction. Despite its importance, most of the enzymes that synthe- β-(1,3-1,4)-D-xylose backbone (8). Xylans of embryophytes have size xylan have yet to be identified. Xylans have a backbone of a β-1,4–linked xylose backbone. Xylans found in dicots are mostly β-1,4–linked xylose residues with substitutions that include α-(1→2)– restricted to the secondary cell walls, and hence a main component linked glucuronosyl, 4-O-methyl glucuronosyl, and α-1,2- and α-1,3- of wood. -

Aberrant Sialylation in Cancer: Biomarker and Potential Target for Therapeutic Intervention?
cancers Review Aberrant Sialylation in Cancer: Biomarker and Potential Target for Therapeutic Intervention? Silvia Pietrobono * and Barbara Stecca * Tumor Cell Biology Unit, Core Research Laboratory, Institute for Cancer Research and Prevention (ISPRO), Viale Pieraccini 6, 50139 Florence, Italy * Correspondence: [email protected] (S.P.); [email protected] (B.S.); Tel.: +39-055-7944568 (S.P.); +39-055-7944567 (B.S.) Simple Summary: Sialylation is a post-translational modification that consists in the addition of sialic acid to growing glycan chains on glycoproteins and glycolipids. Aberrant sialylation is an established hallmark of several types of cancer, including breast, ovarian, pancreatic, prostate, colorectal and lung cancers, melanoma and hepatocellular carcinoma. Hypersialylation can be the effect of increased activity of sialyltransferases and results in an excess of negatively charged sialic acid on the surface of cancer cells. Sialic acid accumulation contributes to tumor progression by several paths, including stimulation of tumor invasion and migration, and enhancing immune evasion and tumor cell survival. In this review we explore the mechanisms by which sialyltransferases promote cancer progression. In addition, we provide insights into the possible use of sialyltransferases as biomarkers for cancer and summarize findings on the development of sialyltransferase inhibitors as potential anti-cancer treatments. Abstract: Sialylation is an integral part of cellular function, governing many biological processes Citation: Pietrobono, S.; Stecca, B. including cellular recognition, adhesion, molecular trafficking, signal transduction and endocytosis. Aberrant Sialylation in Cancer: Sialylation is controlled by the levels and the activities of sialyltransferases on glycoproteins and Biomarker and Potential Target for lipids. Altered gene expression of these enzymes in cancer yields to cancer-specific alterations of Therapeutic Intervention? Cancers glycoprotein sialylation. -

Open Matthew R Moreau Ph.D. Dissertation Finalfinal.Pdf
The Pennsylvania State University The Graduate School Department of Veterinary and Biomedical Sciences Pathobiology Program PATHOGENOMICS AND SOURCE DYNAMICS OF SALMONELLA ENTERICA SEROVAR ENTERITIDIS A Dissertation in Pathobiology by Matthew Raymond Moreau 2015 Matthew R. Moreau Submitted in Partial Fulfillment of the Requirements for the Degree of Doctor of Philosophy May 2015 The Dissertation of Matthew R. Moreau was reviewed and approved* by the following: Subhashinie Kariyawasam Associate Professor, Veterinary and Biomedical Sciences Dissertation Adviser Co-Chair of Committee Bhushan M. Jayarao Professor, Veterinary and Biomedical Sciences Dissertation Adviser Co-Chair of Committee Mary J. Kennett Professor, Veterinary and Biomedical Sciences Vijay Kumar Assistant Professor, Department of Nutritional Sciences Anthony Schmitt Associate Professor, Veterinary and Biomedical Sciences Head of the Pathobiology Graduate Program *Signatures are on file in the Graduate School iii ABSTRACT Salmonella enterica serovar Enteritidis (SE) is one of the most frequent common causes of morbidity and mortality in humans due to consumption of contaminated eggs and egg products. The association between egg contamination and foodborne outbreaks of SE suggests egg derived SE might be more adept to cause human illness than SE from other sources. Therefore, there is a need to understand the molecular mechanisms underlying the ability of egg- derived SE to colonize the chicken intestinal and reproductive tracts and cause disease in the human host. To this end, the present study was carried out in three objectives. The first objective was to sequence two egg-derived SE isolates belonging to the PFGE type JEGX01.0004 to identify the genes that might be involved in SE colonization and/or pathogenesis. -

Arnt Proteins That Catalyse the Glycosylation of Lipopolysaccharide Share Common Features with Bacterial N-Oligosaccharyltransferases
ArnT proteins that catalyse the glycosylation of lipopolysaccharide share common features with bacterial N-oligosaccharyltransferases Tavares-Carreón, F., Mohamed, Y. F., Andrade, A., & Valvano, M. A. (2016). ArnT proteins that catalyse the glycosylation of lipopolysaccharide share common features with bacterial N-oligosaccharyltransferases. Glycobiology, 26(3), 286-300. https://doi.org/10.1093/glycob/cwv095 Published in: Glycobiology Document Version: Peer reviewed version Queen's University Belfast - Research Portal: Link to publication record in Queen's University Belfast Research Portal Publisher rights © [2015] Oxford University Press This is a pre-copyedited, author-produced PDF of an article accepted for publication in Glycobiology following peer review. The version of record Tavares-Carreón, F, Mohamed, YF, Andrade, A & Valvano, MA 2016, 'ArnT proteins that catalyse the glycosylation of lipopolysaccharide share common features with bacterial N-oligosaccharyltransferases' Glycobiology, vol 26, no. 3, pp. 286-300. is available online at:http://glycob.oxfordjournals.org/content/26/3/286 General rights Copyright for the publications made accessible via the Queen's University Belfast Research Portal is retained by the author(s) and / or other copyright owners and it is a condition of accessing these publications that users recognise and abide by the legal requirements associated with these rights. Take down policy The Research Portal is Queen's institutional repository that provides access to Queen's research output. Every effort has been made to ensure that content in the Research Portal does not infringe any person's rights, or applicable UK laws. If you discover content in the Research Portal that you believe breaches copyright or violates any law, please contact [email protected]. -

Fucosyltransferase 8 As a Functional Regulator of Nonsmall Cell Lung Cancer
Fucosyltransferase 8 as a functional regulator of nonsmall cell lung cancer Chien-Yu Chena,b, Yi-Hua Janb, Yi-Hsiu Juanc, Chih-Jen Yangd, Ming-Shyan Huangd, Chong-Jen Yuc, Pan-Chyr Yangc, Michael Hsiaob, Tsui-Ling Hsub,1, and Chi-Huey Wongb,1 aInstitute of Biochemical Sciences, National Taiwan University, Taipei 106, Taiwan; bGenomics Research Center, Academia Sinica, Taipei 115, Taiwan; cDepartment of Internal Medicine, National Taiwan University Hospital, Taipei 100, Taiwan; and dDepartment of Internal Medicine, Kaohsiung Medical University Hospital, Kaohsiung Medical University, Kaohsiung 807, Taiwan Contributed by Chi-Huey Wong, November 26, 2012 (sent for review August 17, 2012) The up-regulation of fucosyltransferase 8 (FUT8), the only enzyme downstream signaling. Furthermore, the increase in core fuco- catalyzing α1,6-fucosylation in mammals, has been observed in sylation on E-cadherin has been shown to strengthen cell–cell several malignant cancers including liver, ovarian, thyroid, and co- adhesion (10). lorectal cancers. However, the pathological role and the regulatory Both transgenic and knockout mouse models have been gen- mechanism of FUT8 in cancers remain largely unknown. In the cur- erated to study the physiological role of FUT8 (7, 11, 12). Ec- rent study, we report that the expression of FUT8 is up-regulated in topic expression of FUT8 in mice results in an accumulation of nonsmall cell lung cancer (NSCLC) and correlates with tumor me- lipid droplets in hepatocytes and proximal renal tubular cells. tastasis, disease recurrence, and poor survival in patients with This steatosis-like phenotype observed in transgenic mice is NSCLC. Knocking down FUT8 in aggressive lung cancer cell lines linked to the activity of liver lysosomal acid lipase, which becomes significantly inhibits their malignant behaviors including in vitro inactive when over core-fucosylated (11), suggesting that excess invasion and cell proliferation, as well as in vivo metastasis and core fucosylation may lead to a breakdown of normal lipid me- tumor growth. -

Estudi Cinètic De L'activitat Enzimàtica De GT-MG517
PART EXPERIMENTAL - Materials i mètodes – - 229 - - 230 - - 231 - - 232 - Capítol 8: Protocols de biologia molecular - 233 - - 234 - Capítol 8: Protocols de biologia molecular 8 Protocols de biologia molecular 8.1 Capítol 2: Estudi de l’essencialitat de les possibles glicosiltransferases de Mycoplasma genitalium 8.1.1 Preparació de medi SP4 La preparació de medi SP4, medi complex que s’usa per al creixement de Mycoplasma genitalium, té lloc com s’explica a continuació. A la Taula 8.1 se’n presenten els components i els volums corresponents per a la preparació de 500 mL de medi líquid. Taula 8.1. Composició i preparació del medi SP4. Quantitat per Compost a 500 mL Preparació de la base PPLO (Difco) 1.75 g Triptona (Difco) 5 g Bactopeptona (Difco) 2.65 g Glucosa (Sigma-Aldrich) 2.5 g Aigua destil·lada Fins a 312 mL Preparació dels complements Yestolate 2 % autoclavat (Difco) 50 mL Roig de fenol 0.1 % pH 7 autoclavat 7 mL Extracte de llevat fresc 25 % 17.5 mL CMRL 10x (Invitrogen) 25 mL Sèrum boví fetal (Invitrogen) 85 mL Glutamina 29.2 mg/mL 1.71 mL Inicialment es mesclen tots els components corresponents a la base del medi i s’ajusta el pH a 7.8 amb NaOH 1 M. El conjunt s’autoclava 15 min a 120 ºC i es deixa refredar abans d’afegir-hi els complements. Un cop aquests s’han addicionat, cal comprovar que el pH del medi es manté entre 7.6 i 7.8. La preparació de l’extracte de llevat té lloc a partir de la dilució de 250 g de llevat fresc en 1 L d’aigua destil·lada. -

Two Arabidopsis Proteins Synthesize Acetylated Xylan Invitro
The Plant Journal (2014) 80, 197–206 doi: 10.1111/tpj.12643 FEATURED ARTICLE Two Arabidopsis proteins synthesize acetylated xylan in vitro Breeanna R. Urbanowicz, Maria J. Pena*,~ Heather A. Moniz, Kelley W. Moremen and William S. York* Complex Carbohydrate Research Center, University of Georgia, 315 Riverbend Road, Athens, GA 30602, USA Received 4 June 2014; revised 18 July 2014; accepted 1 August 2014; published online 21 August 2014. *For correspondence (e-mails [email protected]; [email protected]). SUMMARY Xylan is the third most abundant glycopolymer on earth after cellulose and chitin. As a major component of wood, grain and forage, this natural biopolymer has far-reaching impacts on human life. This highly acetylated cell wall polysaccharide is a vital component of the plant cell wall, which functions as a molecular scaffold, pro- viding plants with mechanical strength and flexibility. Mutations that impair synthesis of the xylan backbone give rise to plants that fail to grow normally because of collapsed xylem cells in the vascular system. Phenotypic analysis of these mutants has implicated many proteins in xylan biosynthesis; however, the enzymes directly responsible for elongation and acetylation of the xylan backbone have not been unambiguously identified. Here we provide direct biochemical evidence that two Arabidopsis thaliana proteins, IRREGULAR XYLEM 10–L (IRX10-L) and ESKIMO1/TRICOME BIREFRINGENCE 29 (ESK1/TBL29), catalyze these respective processes in vi- tro. By identifying the elusive xylan synthase and establishing ESK1/TBL29 as the archetypal plant polysaccha- ride O-acetyltransferase, we have resolved two long-standing questions in plant cell wall biochemistry. -

Multiplexed Engineering Glycosyltransferase Genes in CHO Cells Via Targeted Integration for Producing Antibodies with Diverse Complex‑Type N‑Glycans Ngan T
www.nature.com/scientificreports OPEN Multiplexed engineering glycosyltransferase genes in CHO cells via targeted integration for producing antibodies with diverse complex‑type N‑glycans Ngan T. B. Nguyen, Jianer Lin, Shi Jie Tay, Mariati, Jessna Yeo, Terry Nguyen‑Khuong & Yuansheng Yang* Therapeutic antibodies are decorated with complex‑type N‑glycans that signifcantly afect their biodistribution and bioactivity. The N‑glycan structures on antibodies are incompletely processed in wild‑type CHO cells due to their limited glycosylation capacity. To improve N‑glycan processing, glycosyltransferase genes have been traditionally overexpressed in CHO cells to engineer the cellular N‑glycosylation pathway by using random integration, which is often associated with large clonal variations in gene expression levels. In order to minimize the clonal variations, we used recombinase‑mediated‑cassette‑exchange (RMCE) technology to overexpress a panel of 42 human glycosyltransferase genes to screen their impact on antibody N‑linked glycosylation. The bottlenecks in the N‑glycosylation pathway were identifed and then released by overexpressing single or multiple critical genes. Overexpressing B4GalT1 gene alone in the CHO cells produced antibodies with more than 80% galactosylated bi‑antennary N‑glycans. Combinatorial overexpression of B4GalT1 and ST6Gal1 produced antibodies containing more than 70% sialylated bi‑antennary N‑glycans. In addition, antibodies with various tri‑antennary N‑glycans were obtained for the frst time by overexpressing MGAT5 alone or in combination with B4GalT1 and ST6Gal1. The various N‑glycan structures and the method for producing them in this work provide opportunities to study the glycan structure‑and‑function and develop novel recombinant antibodies for addressing diferent therapeutic applications. -

Induced Structural Changes in a Multifunctional Sialyltransferase
Biochemistry 2006, 45, 2139-2148 2139 Cytidine 5′-Monophosphate (CMP)-Induced Structural Changes in a Multifunctional Sialyltransferase from Pasteurella multocida†,‡ Lisheng Ni,§ Mingchi Sun,§ Hai Yu,§ Harshal Chokhawala,§ Xi Chen,*,§ and Andrew J. Fisher*,§,| Department of Chemistry and the Section of Molecular and Cellular Biology, UniVersity of California, One Shields AVenue, DaVis, California 95616 ReceiVed NoVember 23, 2005; ReVised Manuscript ReceiVed December 19, 2005 ABSTRACT: Sialyltransferases catalyze reactions that transfer a sialic acid from CMP-sialic acid to an acceptor (a structure terminated with galactose, N-acetylgalactosamine, or sialic acid). They are key enzymes that catalyze the synthesis of sialic acid-containing oligosaccharides, polysaccharides, and glycoconjugates that play pivotal roles in many critical physiological and pathological processes. The structures of a truncated multifunctional Pasteurella multocida sialyltransferase (∆24PmST1), in the absence and presence of CMP, have been determined by X-ray crystallography at 1.65 and 2.0 Å resolutions, respectively. The ∆24PmST1 exists as a monomer in solution and in crystals. Different from the reported crystal structure of a bifunctional sialyltransferase CstII that has only one Rossmann domain, the overall structure of the ∆24PmST1 consists of two separate Rossmann nucleotide-binding domains. The ∆24PmST1 structure, thus, represents the first sialyltransferase structure that belongs to the glycosyltransferase-B (GT-B) structural group. Unlike all other known GT-B structures, however, there is no C-terminal extension that interacts with the N-terminal domain in the ∆24PmST1 structure. The CMP binding site is located in the deep cleft between the two Rossmann domains. Nevertheless, the CMP only forms interactions with residues in the C-terminal domain. -
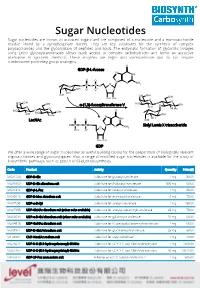
Sugar Nucleotides Sugar Nucleotides Are Known As Activated Sugars and Are Composed of a Nucleoside and a Monosaccharide Residue Linked by a Pyrophosphate Moiety
Sugar Nucleotides Sugar nucleotides are known as activated sugars and are composed of a nucleoside and a monosaccharide residue linked by a pyrophosphate moiety. They are key substrates for the synthesis of complex polysaccharides and the glycosylation of peptides and lipids. The enzymatic formation of glycosidic linkages using Leloir glycosyltransferases allows quick access to complex carbohydrates and forms an attractive alternative to synthetic methods. These enzymes are regio- and stereoselective and do not require cumbersome protecting group strategies. GDP-β-L-fucose α-(1,3)-fucosyltransferase V LacNAc Sialyl Lewis X trisaccharide GDP We offer a wide range of sugar nucleotides as useful building blocks for the preparation of biologically relevant oligosaccharides and glycoconjugates. Also, a range of modified sugar nucleotides is available for the study of biosynthetic pathways, such as Lipid A or (Sia)LeX biosynthesis. Code Product Activity Quantity Price ($) MG31129 GDP-D-Glc Substrate for glucosyltransferase 1 mg 50.00 MU08960 UDP-D-Glc disodium salt Substrate for β-glucosyltransferase 500 mg 60.00 MG01912 GDP-β-L-Fuc Substrate for fucosyltransferase 1 mg 85.00 MG05610 GDP-D-Man disodium salt Substrate for mannosyltransferase 5 mg 75.00 MU07658 UDP-α-D-Xyl Substrate for xylosyltransferase 1 mg 150.00 MU07955 UDP-GlcNAc disodium salt (other salts available) Substrate for acetylglucosaminyltransferase 25 mg 75.00 MU06699 UDP-α-D-Gal disodium salt (other salts available) Substrate for galactosyltransferase 10 mg 68.30 MU04515 UDP-GalNAc