Botany UG 2Nd Semester (Hons) Paper- C4T, Unit- 6- Gymnosperms Anusree Saha Assistant Professor (Department of Botany) Gymnosperms
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Gymnosperms the MESOZOIC: ERA of GYMNOSPERM DOMINANCE
Chapter 24 Gymnosperms THE MESOZOIC: ERA OF GYMNOSPERM DOMINANCE THE VASCULAR SYSTEM OF GYMNOSPERMS CYCADS GINKGO CONIFERS Pinaceae Include the Pines, Firs, and Spruces Cupressaceae Include the Junipers, Cypresses, and Redwoods Taxaceae Include the Yews, but Plum Yews Belong to Cephalotaxaceae Podocarpaceae and Araucariaceae Are Largely Southern Hemisphere Conifers THE LIFE CYCLE OF PINUS, A REPRESENTATIVE GYMNOSPERM Pollen and Ovules Are Produced in Different Kinds of Structures Pollination Replaces the Need for Free Water Fertilization Leads to Seed Formation GNETOPHYTES GYMNOSPERMS: SEEDS, POLLEN, AND WOOD THE ECOLOGICAL AND ECONOMIC IMPORTANCE OF GYMNOSPERMS The Origin of Seeds, Pollen, and Wood Seeds and Pollen Are Key Reproductive SUMMARY Innovations for Life on Land Seed Plants Have Distinctive Vegetative PLANTS, PEOPLE, AND THE Features ENVIRONMENT: The California Coast Relationships among Gymnosperms Redwood Forest 1 KEY CONCEPTS 1. The evolution of seeds, pollen, and wood freed plants from the need for water during reproduction, allowed for more effective dispersal of sperm, increased parental investment in the next generation and allowed for greater size and strength. 2. Seed plants originated in the Devonian period from a group called the progymnosperms, which possessed wood and heterospory, but reproduced by releasing spores. Currently, five lineages of seed plants survive--the flowering plants plus four groups of gymnosperms: cycads, Ginkgo, conifers, and gnetophytes. Conifers are the best known and most economically important group, including pines, firs, spruces, hemlocks, redwoods, cedars, cypress, yews, and several Southern Hemisphere genera. 3. The pine life cycle is heterosporous. Pollen strobili are small and seasonal. Each sporophyll has two microsporangia, in which microspores are formed and divide into immature male gametophytes while still retained in the microsporangia. -
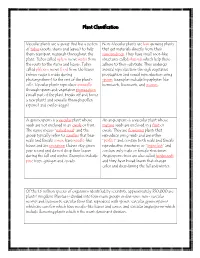
Plant Classification
Plant Classification Vascular plants are a group that has a system Non-Vascular plants are low growing plants of tubes (roots, stems and leaves) to help that get materials directly from their them transport materials throughout the surroundings. They have small root-like plant. Tubes called xylem move water from structures called rhizoids which help them the roots to the stems and leaves. Tubes adhere to their substrate. They undergo called phloem move food from the leaves asexual reproduction through vegetative (where sugar is made during propagation and sexual reproduction using photosynthesis) to the rest of the plant’s spores. Examples include bryophytes like cells. Vascular plants reproduce asexually hornworts, liverworts, and mosses. through spores and vegetative propagation (small part of the plant breaks off and forms a new plant) and sexually through pollen (sperm) and ovules (eggs). A gymnosperm is a vascular plant whose An angiosperm is a vascular plant whose seeds are not enclosed in an ovule or fruit. mature seeds are enclosed in a fruit or The name means “naked seed” and the ovule. They are flowering plants that group typically refers to conifers that bear reproduce using seeds and are either male and female cones, have needle-like “perfect” and contain both male and female leaves and are evergreen (leaves stay green reproductive structures or “imperfect” and year round and do not drop their leaves contain only male or female structures. during the fall and winter. Examples include Angiosperm trees are also called hardwoods pine trees, ginkgos and cycads. and they have broad leaves that change color and drop during the fall and winter. -

B.Sc Botany (Sub.) I Group: a General Characters of Gymnosperm
1 B.Sc Botany (Sub.) I Group: A General Characters of Gymnosperm Gymnosperms are a small group of plants comprising only 70 genera and 725 living species. The word Gymnosperm was used by the Greek botanists Theophrastus in 300 B.C. for the plants with unprotected seeds. Gymnosperms are naked seeded plants. The gymnosperms are characterized by the following features: i. It shows the distinct alternation of generations. ii. Sporophytic generation is the dormant phase of the life cycle. The main plants are sporophyte and the gametophytes are dependent on it throughout. iii. The sporophytic plants are usually tall, woody, perennial trees or shrubs and differentiated into root, stem and leaves. iv. Root- Root is usually well developed tap root system. The stele in root is diarch or polyarch. v. Stem- stems are usually branched but unbranched in Cycas. vi. The leaves may be compound as in Cycas or simple as in Pinus. Internal structure: i. The vascular bundles in stem are arranged in a ring. The bundles are conjoint, collateral and open. ii. The secondary growth is effected by a cambial layer which produces secondary xylem and secondary phloem on the inner and outer side respectively. iii. The secondary wood forming distinct annual ring. Spring wood: It is made up of layer tracheids with the walls and layer lumen. Autumn wood: It is made up of compactly arranged smaller tracheids with thick walls and smaller lumen. iv. The wood may be manoxylic or pycnoxylic and may be monoxylic or polyxylic. Dr. Sanjeev Kumar Vidyarthi, dept. of Botany, Dr. L.K.V.D. -

Life and Time of Indian Williamsonia
Life and time of Indian Williamsonia )ayasri Banerji Banerji, J 1992. Life and time of Indian Williamsonia. Palaeobotanist 40 : 245-259. The Williamsonia plant, belonging to the order Bennettitales, consists of stem-Bucklandia Presl, leaf Ptilophyllum Morris, male flower- Weltrichia Braun and female flower- Williamsonia Carruthers. This plant was perhaps a small, much branched woody tree of xerophytic environment. It co-existed alongwith extremely variable and rich flora including highly diversified plant groups from algae to gymnosperms. In India, it appeared during the marine Jurassic, proliferated and widely distributed in the Lower Cretaceous and disappeared from the vegetational scenario of Upper Cretaceous Period with the advent of angiosperms. Key-words-Bennettitales, Williamsonia, Jurassic-Cretaceous (India). jayasri Baner}i, Birbal Sahni Institute of Palaeobotany, 53 University Road, Lucknow 226007, India. ri~T ~ 'lfmftq- fili'<1QQ«lf.:lQi "" om~~ "l?li'O'$i'OI<1fl \f>'f it ~ filf<1QQ«lf.:lQi qtfr if ~ ~ 3!<'flT-3!<'flT 'lTIif it ~ "lTif t W'I'T CAT-~ m, ~ ~ ~it ~fu;m;;mrr~1 ~qtm~~ <ffir ll~<'ilf4>~<'1'i"li'tfGr, "''!'''l ~~?l~Ti:t>Qi <ilf.1 'I"lT filf<1QQ«lf.:lQi ~ ~ ~ f;;ru-if.~ ~ 61'1'~dOl'h\'i if m <'IT<'1T ~ Wc:r, 3!fuq,iffiliOlT3if it ~ ¥i "IT' ~ it il'1f4ff1"1ld, it if qtfr ~ 'it, q;r tt ~ ~tl W'I'T~~'-I'<"1if~~3lT, 3Tahmm'-l'<"1if~~~-~'d"f~<f"lT'3'1f'<mm~ ~ ~ ~ ~ if qtm if if t1T"f -flT"f lIT lfllT' LIFE OF WILLIAMSONIA PLANT B. dichotoma Sharma. In B. indica Seward, the secondary wood is more compact than recent cycads In the Upper Mesozoic Era, a new group of and cycadeoids. -

Dr. Sahanaj Jamil Associate Professor of Botany M.L.S.M. College, Darbhanga
Subject BOTANY Paper No V Paper Code BOT521 Topic Taxonomy and Diversity of Seed Plant: Gymnosperms & Angiosperms Dr. Sahanaj Jamil Associate Professor of Botany M.L.S.M. College, Darbhanga BOTANY PG SEMESTER – II, PAPER –V BOT521: Taxonomy and Diversity of seed plants UNIT- I BOTANY PG SEMESTER – II, PAPER –V BOT521: Taxonomy and Diversity of seed plants Classification of Gymnosperms. # Robert Brown (1827) for the first time recognized Gymnosperm as a group distinct from angiosperm due to the presence of naked ovules. BENTHAM and HOOKSER (1862-1883) consider them equivalent to dicotyledons and monocotyledons and placed between these two groups of angiosperm. They recognized three classes of gymnosperm, Cyacadaceae, coniferac and gnetaceae. Later ENGLER (1889) created a group Gnikgoales to accommodate the genus giankgo. Van Tieghem (1898) treated Gymnosperm as one of the two subdivision of spermatophyte. To accommodate the fossil members three more classes- Pteridospermae, Cordaitales, and Bennettitales where created. Coulter and chamberlain (1919), Engler and Prantl (1926), Rendle (1926) and other considered Gymnosperm as a division of spermatophyta, Phanerogamia or Embryoptyta and they further divided them into seven orders: - i) Cycadofilicales ii) Cycadales iii) Bennettitales iv) Ginkgoales v) Coniferales vi) Corditales vii) Gnetales On the basis of wood structure steward (1919) divided Gymnosperm into two classes: - i) Manoxylic ii) Pycnoxylic The various classification of Gymnosperm proposed by various workers are as follows: - i) Sahni (1920): - He recognized two sub-divison in gymnosperm: - a) Phylospermae b) Stachyospermae BOTANY PG SEMESTER – II, PAPER –V BOT521: Taxonomy and Diversity of seed plants ii) Classification proposed by chamber lain (1934): - He divided Gymnosperm into two divisions: - a) Cycadophyta b) Coniterophyta iii) Classification proposed by Tippo (1942):- He considered Gymnosperm as a class of the sub- phylum pteropsida and divided them into two sub classes:- a) Cycadophyta b) Coniferophyta iv) D. -

Ecological Sorting of Vascular Plant Classes During the Paleozoic Evolutionary Radiation
i1 Ecological Sorting of Vascular Plant Classes During the Paleozoic Evolutionary Radiation William A. DiMichele, William E. Stein, and Richard M. Bateman DiMichele, W.A., Stein, W.E., and Bateman, R.M. 2001. Ecological sorting of vascular plant classes during the Paleozoic evolutionary radiation. In: W.D. Allmon and D.J. Bottjer, eds. Evolutionary Paleoecology: The Ecological Context of Macroevolutionary Change. Columbia University Press, New York. pp. 285-335 THE DISTINCTIVE BODY PLANS of vascular plants (lycopsids, ferns, sphenopsids, seed plants), corresponding roughly to traditional Linnean classes, originated in a radiation that began in the late Middle Devonian and ended in the Early Carboniferous. This relatively brief radiation followed a long period in the Silurian and Early Devonian during wrhich morphological complexity accrued slowly and preceded evolutionary diversifications con- fined within major body-plan themes during the Carboniferous. During the Middle Devonian-Early Carboniferous morphological radiation, the major class-level clades also became differentiated ecologically: Lycopsids were cen- tered in wetlands, seed plants in terra firma environments, sphenopsids in aggradational habitats, and ferns in disturbed environments. The strong con- gruence of phylogenetic pattern, morphological differentiation, and clade- level ecological distributions characterizes plant ecological and evolutionary dynamics throughout much of the late Paleozoic. In this study, we explore the phylogenetic relationships and realized ecomorphospace of reconstructed whole plants (or composite whole plants), representing each of the major body-plan clades, and examine the degree of overlap of these patterns with each other and with patterns of environmental distribution. We conclude that 285 286 EVOLUTIONARY PALEOECOLOGY ecological incumbency was a major factor circumscribing and channeling the course of early diversification events: events that profoundly affected the structure and composition of modern plant communities. -

A Global Analysis of the Distribution and Conservation Status Of
Journal of Biogeography (J. Biogeogr.) (2015) 42, 809–820 SYNTHESIS Fighting their last stand? A global analysis of the distribution and conservation status of gymnosperms Yann Fragniere1,Sebastien Betrisey2,3,Leonard Cardinaux1, Markus Stoffel4,5 and Gregor Kozlowski1,2* 1Natural History Museum Fribourg, CH-1700 ABSTRACT Fribourg, Switzerland, 2Department of Biology Aim Gymnosperms are often described as a marginal and threatened group, and Botanic Garden, University of Fribourg, members of which tend to be out-competed by angiosperms and which therefore CH-1700 Fribourg, Switzerland, 3Conservation Biogeography, Department of preferentially persist at higher latitudes and elevations. The aim of our synthesis Geosciences, University of Fribourg, CH-1700 was to test these statements by investigating the global latitudinal and elevational Fribourg, Switzerland, 4Dendrolab.ch, distribution of gymnosperms, as well as their conservation status, using all extant Institute of Geological Sciences, University of gymnosperm groups (cycads, gnetophytes, ginkgophytes and conifers). 5 Bern, CH-3012 Bern, Switzerland, Institute Location Worldwide. for Environmental Sciences, Climatic Change and Climate Impacts, University of Geneva, Methods We developed a database of 1014 species of gymnosperms containing CH-1227 Carouge, Switzerland latitudinal and elevational distribution data, as well as their global conservation status, as described in the literature. The 1014 species comprised 305 cycads, 101 gnetophytes, the only living representative of ginkgophytes, and 607 conifers. Generalized additive models, frequency histograms, kernel density estimations and distribution maps based on Takhtajan’s floristic regions were used. Results Although the diversity of gymnosperms decreases at equatorial lati- tudes, approximately 50% of the extant species occur primarily between the tropics. More than 43% of gymnosperms can occur at very low elevations (≤ 200 m a.s.l.). -

Name That Gymnosperm
A B C D This tree is found frequently This evergreen is found throughout This species is found at drier and This Wyoming tree is a remnant of throughout the eastern half of the state. The species is known for lower elevations compared to some the last ice age and is found exclu- Wyoming ranging from the Black often having serotinous cones that of the other trees pictured. This tree sively in the Black Hills of Wyoming Hills to the Laramie Mountains and only open to release seeds when the shares its name with the town in and South Dakota. It grows at higher the Bighorns. The tree is known cone is heated. Once the limbs are central Wyoming. elevations along riparian or wet areas for its ability to withstand the heat removed, this tree makes an excel- in its native habitat. of wildfires because of its thick, lent pole for teepees because of its reddish-colored bark. long, slender trunk. NAME THAT GYMNOSPERM Wyoming is host to many conifer (gymnosperm or “naked in hot, dry, and low elevations while others are found in cold and seed” plants including conifer, cycads, and ginkos) tree species. high elevations. Cones are an excellent tool that can be used for The cones in this quiz and their parent trees are found at different identification of evergreens. Match the tree to the photo. Good geographical locations across the Cowboy State. Some are found luck and keep an eye out for these trees this year! E F G H Known for having extremely flexible This species is probably better known This tree is found in most high- This tree grows at high elevations limbs, this tree is often found brav- as an ornamental in Wyoming but is elevation forests of Wyoming. -

CPY Document
CHAPTER 1 Plant diversity has evolved and is evolving on the earth today according to the processes of natural selection. All basic "body plans" of vascular plants were in existence by the Early Carboniferous, approximately 340 million years ago, and each major radiation occupied a particular habitat type, for example, wetland versus terra firma, pristine versus disturbed environments. Today the great spe- cies diversity we see on theplanetis dominated by just a few of these hasicplant types. This chapter sets the stage to understanding the origin and radiation of plant diversity on the earth before the significant influence of humans. The im- plication is that plants may become more diverse in the number of species level over time, but less diverse in overall form. The history of life tells us that plants that are dominant and diverse today will certainly be inconspicuous or even ex--' tinct in the distant future. EVOLUTION OF LAND PLANT DIVERSITY MAJOR INNOVATIONS AND LINEAGES THROUGH TIME William A. DiMichele and Richard M. Bateman PLANT DIVERSITY VIEWED through the lens of deep time takes on a con- siderably different aspect than when examined in the present. Although the fossil record captures only fragments of the terrestrial world of the past 450 million years, it does indicate clearly that the world of today is simply a passing phase, the latest permutation in a string of spasmodic changes in the ecological organization of the terrestrial biosphere. The pace of change in species diversity and that of ecosystem structure and composition have followed broadly parallel paths, unquestionably related but not always changing in unison. -

Curriculum Vitae
CURRICULUM VITAE ORCID ID: 0000-0003-0186-6546 Gar W. Rothwell Edwin and Ruth Kennedy Distinguished Professor Emeritus Department of Environmental and Plant Biology Porter Hall 401E T: 740 593 1129 Ohio University F: 740 593 1130 Athens, OH 45701 E: [email protected] also Courtesy Professor Department of Botany and PlantPathology Oregon State University T: 541 737- 5252 Corvallis, OR 97331 E: [email protected] Education Ph.D.,1973 University of Alberta (Botany) M.S., 1969 University of Illinois, Chicago (Biology) B.A., 1966 Central Washington University (Biology) Academic Awards and Honors 2018 International Organisation of Palaeobotany lifetime Honorary Membership 2014 Fellow of the Paleontological Society 2009 Distinguished Fellow of the Botanical Society of America 2004 Ohio University Distinguished Professor 2002 Michael A. Cichan Award, Botanical Society of America 1999-2004 Ohio University Presidential Research Scholar in Biomedical and Life Sciences 1993 Edgar T. Wherry Award, Botanical Society of America 1991-1992 Outstanding Graduate Faculty Award, Ohio University 1982-1983 Chairman, Paleobotanical Section, Botanical Society of America 1972-1973 University of Alberta Dissertation Fellow 1971 Paleobotanical (Isabel Cookson) Award, Botanical Society of America Positions Held 2011-present Courtesy Professor of Botany and Plant Pathology, Oregon State University 2008-2009 Visiting Senior Researcher, University of Alberta 2004-present Edwin and Ruth Kennedy Distinguished Professor of Environmental and Plant Biology, Ohio -

Revision of the Talbragar Fish Bed Flor (Jurassic)
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS White, Mary E., 1981. Revision of the Talbragar Fish Bed Flora (Jurassic) of New South Wales. Records of the Australian Museum 33(15): 695–721. [31 July 1981]. doi:10.3853/j.0067-1975.33.1981.269 ISSN 0067-1975 Published by the Australian Museum, Sydney naturenature cultureculture discover discover AustralianAustralian Museum Museum science science is is freely freely accessible accessible online online at at www.australianmuseum.net.au/publications/www.australianmuseum.net.au/publications/ 66 CollegeCollege Street,Street, SydneySydney NSWNSW 2010,2010, AustraliaAustralia REVISION OF THE TALBRAGAR FISH BED FLORA (jURASSiC) OF NEW SOUTH WALES MARY E. WH ITE The Australian Museum, Sydney. SUMMARY The three well known form-species of the Talbragar Fish Bed Flora-Podozamites lanceolatus, Elatocladus planus and Taeniopteris spa tu lata - are redescribed as Agathis jurassica sp. nov., Rissikia talbragarensis sp. novo and Pentoxylon australica sp. novo respectively. The minor components of the assemblage are described and illustrated, and in some cases, reclassified. Additions are made to the list of plants recorded from the horizon. INTRODUCTION The Talbragar Fish Beds are characterised by their beautifully preserved fish and plant remains which occur in great profusion throughout the shale lens which comprises the Beds. The ochre-coloured shale is ferruginous, with impressions of plants and fish, white in colour, standing out dramatically. The weathering of the outer layers of blocks of the shale has resulted in contrasting bands of iron-rich stain framing many of the specimens and enhancing their appearance. Specimens are much prized by collectors. The fossil locality is the valley ofthe Talbragar River, about twenty miles due North of Home Rule Mine in the Cassilis District, "on the southern boundary of Boyce's selection" (Anderson 1889). -

Botany Syllabus Semester Pattern with Choice Based Credits System
Shiksha Mandal’s Jankidevi Bajaj College of Science, Wardha. (Autonomous) NAAC (UGC) Reaccredited ‘A’ Institution (A Linguistic Minority College) COLLEGE WITH POTENTIAL FOR EXCELLENCE Star College Status by DBT Govt. of India M.Sc. Botany Syllabus Semester pattern with Choice Based Credits System (2017-2018) Department of Botany Jankidevi Bajaj College of Science, Wardha. JANKIDEVI BAJAJ COLLEGE OF SCIENCE, WARDHA Two Year Post Graduate Course (M. Sc.) SEMESTER PATTERN SYLLABUS (Proposed Under Autonomy) SUBJECT – BOTANY (Distribution of Units) Seme Paper Existing Syllabus Proposed Syllabus ster Uni Content of Unit Alloted Uni Content of Unit Alloted t Hours t Hours No No Seme Paper I I-IV Prokaryotes & 60 I-III Prokaryotes & Viruses, 48 ster I Viruses, Phycology, Mycology Phycology, and Plant Pathology Mycology and IV Microscopy & 12 Plant Pathology Centrifugation Paper II I-IV Bryophytes, 60 I-III Bryophytes, 48 Pteridophytes Pteridophytes IV Plant Microtechniques 12 Paper III I-IV Paleobotany, 60 I-III Paleobotany, 48 Gymnosperms Gymnosperms IV Instrumentation 12 (Spectrophotometery & Chromatography) Paper IV I-IV Cytology, 60 I-III Cytology, Genetics 48 Genetics IV Methods To Study Cell 12 / Tissue Structure Seme Paper V I-IV Plant Physiology, 60 I-III Plant Physiology, 48 ster II Biochemistry Biochemistry IV Analytical 12 Pharmacognosy Paper VI I-IV Plant 60 I-III Plant Development, 48 Development, Reproduction Reproduction IV Phytochemistry 12 Paper VII I-IV Cell, Molecular 60 I-III Cell, Molecular 48 Biology- I Biology- I IV Data