The Origin of Monocotyledons from Dicotyledons, Through Self-Adaptation to a Moist
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Diversity and Distribution of Family Araceae in Doi Inthanon National Park, Chiang Mai Province
Agriculture and Natural Resources 52 (2018) 125e131 Contents lists available at ScienceDirect Agriculture and Natural Resources journal homepage: http://www.journals.elsevier.com/agriculture-and- natural-resources/ Original Article Diversity and distribution of family Araceae in Doi Inthanon National Park, Chiang Mai province * Oraphan Sungkajanttranon,a, Dokrak Marod,b Kriangsak Thanompunc a Graduate School, Kasetsart University, Bangkok, 10900, Thailand b Department of Forest Biology, Faculty of Forestry, Kasetsart University, Bangkok, 10900, Thailand c Department of National Parks, Wildlife and Plant Conservation, Bangkok, 10900, Thailand article info abstract Article history: Species of the family Araceae are known for their ethnobotanical utilization; however their species di- Received 28 November 2016 versity is less documented. This study aimed to clarify the relationships between species diversity of the Accepted 18 December 2017 Araceae and altitudinal gradients in the mountain ecosystems in Doi Inthanon National Park, Chiang Mai Available online 17 July 2018 province, northern Thailand. In 2014, tree permanent plots (4 m  4 m) including one strip plot (5 m  500 m) were established every 300 m above mean sea level (amsl) in the range 300e2565 m Keywords: amsl. All species of Araceae were investigated and environmental factors were also recorded. The results Altitudinal gradient showed that 23 species were mostly found in the rainy season and identified into 11 genera: Alocasia, Araceae Doi Inthanon National Park Amorphophallus, Arisaema, Colocasia, Lasia, Pothos, Rhaphidophora, Remusatia, Sauromatum, Scindapsus, fi Ecological niche and Typhonium. The top ve dominant species were Arisaema consanguineum, Amorphophallus fuscus, Species distribution Remusatia hookeriana, Amorphophallus yunnanensis and Colocasia esculenta. Low species diversity was found at the lowest and highest altitude. -

197-1572431971.Pdf
Innovare Journal of Critical Reviews Academic Sciences ISSN- 2394-5125 Vol 2, Issue 2, 2015 Review Article EPIPREMNUM AUREUM (JADE POTHOS): A MULTIPURPOSE PLANT WITH ITS MEDICINAL AND PHARMACOLOGICAL PROPERTIES ANJU MESHRAM, NIDHI SRIVASTAVA* Department of Bioscience and Biotechnology, Banasthali University, Rajasthan, India Email: [email protected] Received: 13 Dec 2014 Revised and Accepted: 10 Jan 2015 ABSTRACT Plants belonging to the Arum family (Araceae) are commonly known as aroids as they contain crystals of calcium oxalate and toxic proteins which can cause intense irritation of the skin and mucous membranes, and poisoning if the raw plant tissue is eaten. Aroids range from tiny floating aquatic plants to forest climbers. Many are cultivated for their ornamental flowers or foliage and others for their food value. Present article critically reviews the growth conditions of Epipremnum aureum (Linden and Andre) Bunting with special emphasis on their ethnomedicinal uses and pharmacological activities, beneficial to both human and the environment. In this article, we review the origin, distribution, brief morphological characters, medicinal and pharmacological properties of Epipremnum aureum, commonly known as ornamental plant having indoor air pollution removing capacity. There are very few reports to the medicinal properties of E. aureum. In our investigation, it has been found that each part of this plant possesses antibacterial, anti-termite and antioxidant properties. However, apart from these it can also turn out to be anti-malarial, anti- cancerous, anti-tuberculosis, anti-arthritis and wound healing etc which are a severe international problem. In the present study, details about the pharmacological actions of medicinal plant E. aureum (Linden and Andre) Bunting and Epipremnum pinnatum (L.) Engl. -

Well-Known Plants in Each Angiosperm Order
Well-known plants in each angiosperm order This list is generally from least evolved (most ancient) to most evolved (most modern). (I’m not sure if this applies for Eudicots; I’m listing them in the same order as APG II.) The first few plants are mostly primitive pond and aquarium plants. Next is Illicium (anise tree) from Austrobaileyales, then the magnoliids (Canellales thru Piperales), then monocots (Acorales through Zingiberales), and finally eudicots (Buxales through Dipsacales). The plants before the eudicots in this list are considered basal angiosperms. This list focuses only on angiosperms and does not look at earlier plants such as mosses, ferns, and conifers. Basal angiosperms – mostly aquatic plants Unplaced in order, placed in Amborellaceae family • Amborella trichopoda – one of the most ancient flowering plants Unplaced in order, placed in Nymphaeaceae family • Water lily • Cabomba (fanwort) • Brasenia (watershield) Ceratophyllales • Hornwort Austrobaileyales • Illicium (anise tree, star anise) Basal angiosperms - magnoliids Canellales • Drimys (winter's bark) • Tasmanian pepper Laurales • Bay laurel • Cinnamon • Avocado • Sassafras • Camphor tree • Calycanthus (sweetshrub, spicebush) • Lindera (spicebush, Benjamin bush) Magnoliales • Custard-apple • Pawpaw • guanábana (soursop) • Sugar-apple or sweetsop • Cherimoya • Magnolia • Tuliptree • Michelia • Nutmeg • Clove Piperales • Black pepper • Kava • Lizard’s tail • Aristolochia (birthwort, pipevine, Dutchman's pipe) • Asarum (wild ginger) Basal angiosperms - monocots Acorales -

Ethnomedicinal Profile of Flora of District Sialkot, Punjab, Pakistan
ISSN: 2717-8161 RESEARCH ARTICLE New Trend Med Sci 2020; 1(2): 65-83. https://dergipark.org.tr/tr/pub/ntms Ethnomedicinal Profile of Flora of District Sialkot, Punjab, Pakistan Fozia Noreen1*, Mishal Choudri2, Shazia Noureen3, Muhammad Adil4, Madeeha Yaqoob4, Asma Kiran4, Fizza Cheema4, Faiza Sajjad4, Usman Muhaq4 1Department of Chemistry, Faculty of Natural Sciences, University of Sialkot, Punjab, Pakistan 2Department of Statistics, Faculty of Natural Sciences, University of Sialkot, Punjab, Pakistan 3Governament Degree College for Women, Malakwal, District Mandi Bahauddin, Punjab, Pakistan 4Department of Chemistry, Faculty of Natural Sciences, University of Gujrat Sialkot Subcampus, Punjab, Pakistan Article History Abstract: An ethnomedicinal profile of 112 species of remedial Received 30 May 2020 herbs, shrubs, and trees of 61 families with significant Accepted 01 June 2020 Published Online 30 Sep 2020 gastrointestinal, antimicrobial, cardiovascular, herpetological, renal, dermatological, hormonal, analgesic and antipyretic applications *Corresponding Author have been explored systematically by circulating semi-structured Fozia Noreen and unstructured questionnaires and open ended interviews from 40- Department of Chemistry, Faculty of Natural Sciences, 74 years old mature local medicine men having considerable University of Sialkot, professional experience of 10-50 years in all the four geographically Punjab, Pakistan diversified subdivisions i.e. Sialkot, Daska, Sambrial and Pasrur of E-mail: [email protected] district Sialkot with a total area of 3106 square kilometres with ORCID:http://orcid.org/0000-0001-6096-2568 population density of 1259/km2, in order to unveil botanical flora for world. Family Fabaceae is found to be the most frequent and dominant family of the region. © 2020 NTMS. -

Phylogeny and Taxonomy of the Genus Elettaria Maton
Cardamoms of South East Asia: phylogeny and taxonomy of the genus Elettaria Maton Helena Båserud Mathisen Master of Science Thesis 2014 Department of Biosciences Faculty of Mathematics and Natural Sciences University of Oslo, Norway © Helena Båserud Mathisen 2014 Cardamoms of South East Asia: phylogeny and taxonomy of the genus Elettaria Illustration on the front page: From White (1811) https://www.duo.uio.no/ Print: Reprosentralen, University of Oslo Acknowledgements There are plenty of people who deserve a big depth of gratitude when I hand in my master thesis today. First of all, I would like to thank my supervisors Axel Dalberg Poulsen, Charlotte Sletten Bjorå and Mark Newman for all help, patience and valuable input over the last 1.5 years, and especially the last couple of weeks. I could not have done this without you guys! Thanks to the approval of our research permit from the Forest Department in Sarawak, Axel and I were able to travel to Borneo and collect plants for my project. I would like to thank the Botanical Research Centre at Semenggoh Wildlife Centre in Sarawak, for all the help we got, and a special thanks goes to Julia, Ling and Vilma for planning and organizing the field trips for us. I would never have mastered the lab technics at Tøyen without good help and guideance from Audun. Thank you for answering my numerous questions so willingly. I would also like to thank My Hanh, Kjersti, Anette and Kine, for inviting me over for dinner and improving my draft and of course my fellow students at the botanical museum (Anne Marte, Karen and Øystein). -

TAXON:Phoenix Sylvestris SCORE:5.0 RATING:Evaluate
TAXON: Phoenix sylvestris SCORE: 5.0 RATING: Evaluate Taxon: Phoenix sylvestris Family: Arecaceae Common Name(s): date sugar palm Synonym(s): Elate sylvestris L. (basionym) Indian date silver date palm wild date palm Assessor: No Assessor Status: Assessor Approved End Date: 29 Jul 2014 WRA Score: 5.0 Designation: EVALUATE Rating: Evaluate Keywords: Naturalized, Tropical Palm, Spiny, Dioecious, Bird-dispersed Qsn # Question Answer Option Answer 101 Is the species highly domesticated? y=-3, n=0 n 102 Has the species become naturalized where grown? 103 Does the species have weedy races? Species suited to tropical or subtropical climate(s) - If 201 island is primarily wet habitat, then substitute "wet (0-low; 1-intermediate; 2-high) (See Appendix 2) High tropical" for "tropical or subtropical" 202 Quality of climate match data (0-low; 1-intermediate; 2-high) (See Appendix 2) High 203 Broad climate suitability (environmental versatility) y=1, n=0 n Native or naturalized in regions with tropical or 204 y=1, n=0 y subtropical climates Does the species have a history of repeated introductions 205 y=-2, ?=-1, n=0 y outside its natural range? 301 Naturalized beyond native range y = 1*multiplier (see Appendix 2), n= question 205 y 302 Garden/amenity/disturbance weed n=0, y = 1*multiplier (see Appendix 2) n 303 Agricultural/forestry/horticultural weed n=0, y = 2*multiplier (see Appendix 2) n 304 Environmental weed n=0, y = 2*multiplier (see Appendix 2) n 305 Congeneric weed n=0, y = 1*multiplier (see Appendix 2) y 401 Produces spines, thorns or burrs -

Outline of Angiosperm Phylogeny
Outline of angiosperm phylogeny: orders, families, and representative genera with emphasis on Oregon native plants Priscilla Spears December 2013 The following listing gives an introduction to the phylogenetic classification of the flowering plants that has emerged in recent decades, and which is based on nucleic acid sequences as well as morphological and developmental data. This listing emphasizes temperate families of the Northern Hemisphere and is meant as an overview with examples of Oregon native plants. It includes many exotic genera that are grown in Oregon as ornamentals plus other plants of interest worldwide. The genera that are Oregon natives are printed in a blue font. Genera that are exotics are shown in black, however genera in blue may also contain non-native species. Names separated by a slash are alternatives or else the nomenclature is in flux. When several genera have the same common name, the names are separated by commas. The order of the family names is from the linear listing of families in the APG III report. For further information, see the references on the last page. Basal Angiosperms (ANITA grade) Amborellales Amborellaceae, sole family, the earliest branch of flowering plants, a shrub native to New Caledonia – Amborella Nymphaeales Hydatellaceae – aquatics from Australasia, previously classified as a grass Cabombaceae (water shield – Brasenia, fanwort – Cabomba) Nymphaeaceae (water lilies – Nymphaea; pond lilies – Nuphar) Austrobaileyales Schisandraceae (wild sarsaparilla, star vine – Schisandra; Japanese -

Medicinal Practices of Sacred Natural Sites: a Socio-Religious Approach for Successful Implementation of Primary
Medicinal practices of sacred natural sites: a socio-religious approach for successful implementation of primary healthcare services Rajasri Ray and Avik Ray Review Correspondence Abstract Rajasri Ray*, Avik Ray Centre for studies in Ethnobiology, Biodiversity and Background: Sacred groves are model systems that Sustainability (CEiBa), Malda - 732103, West have the potential to contribute to rural healthcare Bengal, India owing to their medicinal floral diversity and strong social acceptance. *Corresponding Author: Rajasri Ray; [email protected] Methods: We examined this idea employing ethnomedicinal plants and their application Ethnobotany Research & Applications documented from sacred groves across India. A total 20:34 (2020) of 65 published documents were shortlisted for the Key words: AYUSH; Ethnomedicine; Medicinal plant; preparation of database and statistical analysis. Sacred grove; Spatial fidelity; Tropical diseases Standard ethnobotanical indices and mapping were used to capture the current trend. Background Results: A total of 1247 species from 152 families Human-nature interaction has been long entwined in has been documented for use against eighteen the history of humanity. Apart from deriving natural categories of diseases common in tropical and sub- resources, humans have a deep rooted tradition of tropical landscapes. Though the reported species venerating nature which is extensively observed are clustered around a few widely distributed across continents (Verschuuren 2010). The tradition families, 71% of them are uniquely represented from has attracted attention of researchers and policy- any single biogeographic region. The use of multiple makers for its impact on local ecological and socio- species in treating an ailment, high use value of the economic dynamics. Ethnomedicine that emanated popular plants, and cross-community similarity in from this tradition, deals health issues with nature- disease treatment reflects rich community wisdom to derived resources. -

Understanding the Origin and Rapid Diversification of the Genus Anthurium Schott (Araceae), Integrating Molecular Phylogenetics, Morphology and Fossils
University of Missouri, St. Louis IRL @ UMSL Dissertations UMSL Graduate Works 8-3-2011 Understanding the origin and rapid diversification of the genus Anthurium Schott (Araceae), integrating molecular phylogenetics, morphology and fossils Monica Maria Carlsen University of Missouri-St. Louis, [email protected] Follow this and additional works at: https://irl.umsl.edu/dissertation Part of the Biology Commons Recommended Citation Carlsen, Monica Maria, "Understanding the origin and rapid diversification of the genus Anthurium Schott (Araceae), integrating molecular phylogenetics, morphology and fossils" (2011). Dissertations. 414. https://irl.umsl.edu/dissertation/414 This Dissertation is brought to you for free and open access by the UMSL Graduate Works at IRL @ UMSL. It has been accepted for inclusion in Dissertations by an authorized administrator of IRL @ UMSL. For more information, please contact [email protected]. Mónica M. Carlsen M.S., Biology, University of Missouri - St. Louis, 2003 B.S., Biology, Universidad Central de Venezuela – Caracas, 1998 A Thesis Submitted to The Graduate School at the University of Missouri – St. Louis in partial fulfillment of the requirements for the degree Doctor of Philosophy in Biology with emphasis in Ecology, Evolution and Systematics June 2011 Advisory Committee Peter Stevens, Ph.D. (Advisor) Thomas B. Croat, Ph.D. (Co-advisor) Elizabeth Kellogg, Ph.D. Peter M. Richardson, Ph.D. Simon J. Mayo, Ph.D Copyright, Mónica M. Carlsen, 2011 Understanding the origin and rapid diversification of the genus Anthurium Schott (Araceae), integrating molecular phylogenetics, morphology and fossils Mónica M. Carlsen M.S., Biology, University of Missouri - St. Louis, 2003 B.S., Biology, Universidad Central de Venezuela – Caracas, 1998 Advisory Committee Peter Stevens, Ph.D. -

Arizona Landscape Palms
Cooperative Extension ARIZONA LANDSCAPE PALMS ELIZABETH D AVISON Department of Plant Sciences JOHN BEGEMAN Pima County Cooperative Extension AZ1021 • 12/2000 Issued in furtherance of Cooperative Extension work acts of May 8 and June 30, 1914, in cooperation with the U.S. Department of Agriculture, James A. Christenson, Director, Cooperative Extension, College of Agriculture and Life Sciences, The University of Arizona. The University of Arizona College of Agriculture and Life Sciences is an equal opportunity employer authorized to provide research, educational information and other services to individuals and institutions that function without regard to sex, race, religion, color, national origin, age, Vietnam Era Veteran's status, or disability. Contents Landscape Use ......................................... 3 Adaptation ................................................ 3 Planting Palms ......................................... 3 Care of Established Palms...................... 5 Diseases and Insect Pests ....................... 6 Palms for Arizona .................................... 6 Feather Palms ........................................... 8 Fan Palms................................................ 12 Palm-like Plants ..................................... 16 This information has been reviewed by university faculty. ag.arizona.edu/pubs/garden/az1121.pdf 2 The luxuriant tropical appearance and stately Adaptation silhouette of palms add much to the Arizona landscape. Palms generally can be grown below the 4000 ft level Few other plants are as striking in low and mid elevation in Arizona. However, microclimate may make the gardens. Although winter frosts and low humidity limit difference between success and failure in a given location. the choices somewhat, a good number of palms are Frost pockets, where nighttime cold air tends to collect, available, ranging from the dwarf Mediterranean Fan should be avoided, especially for the tender species. Palms palm to the massive Canary Island Date palm. -
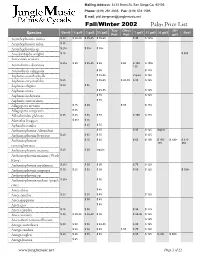
Winter-Fall Sale 2002 Palm Trees-Web
Mailing Address: 3233 Brant St. San Diego Ca, 92103 Phone: (619) 291 4605 Fax: (619) 574 1595 E mail: [email protected] Fall/Winter 2002 Palm Price List Tree Citrus 25/+ Band$ 1 gal$ 2 gal$ 3/5 gal$ 7 gal$ 15 gal$ 20 gal$ Box$ Species Pot$ Pot$ gal$ Acanthophoenix crinita $ 30 $ 30-40 $ 35-45 $ 55-65 $ 95 $ 125+ Acanthophoenix rubra $ 35 Acanthophoenix sp. $ 25+ $ 35+ $ 55+ Acoelorrhaphe wrightii $ 15 $ 300 Acrocomia aculeata $ 25+ $ 35 $ 35-45 $ 65 $ 65 $ 100- $ 150+ Actinokentia divaricata 135 Actinorhytis calapparia $ 55 $ 125 Aiphanes acanthophylla $ 45-55 inquire $ 125 Aiphanes caryotaefolia $ 25 $ 55-65 $ 45-55 $ 85 $ 125 Aiphanes elegans $ 20 $ 35 Aiphanes erosa $ 45-55 $ 125 Aiphanes lindeniana $ 55 $ 125 Aiphanes vincentsiana $ 55 Allagoptera arenaria $ 25 $ 40 $ 55 $ 135 Allagoptera campestris $ 35 Alloschmidtia glabrata $ 35 $ 45 $ 55 $ 85 $ 150 $ 175 Alsmithia longipes $ 35+ $ 55 Aphandra natalia $ 35 $ 55 Archontophoenix Alexandrae $ 55 $ 85 $ 125 inquire Archontophoenix Beatricae $ 20 $ 35 $ 55 $ 125 Archontophoenix $ 25 $ 45 $ 65 $ 100 $ 150- $ 200+ $ 310- 175 350 cunninghamiana Archontophoenix maxima $ 25 $ 30 inquire Archontophoenix maxima (Wash River) Archontophoenix myolaensis $ 25+ $ 30 $ 50 $ 75 $ 125 Archontophoenix purpurea $ 30 $ 25 $ 35 $ 50 $ 85 $ 125 $ 300+ Archontophoenix sp. Archontophoenix tuckerii (peach $ 25+ $ 55 river) Areca alicae $ 45 Areca catechu $ 20 $ 35 $ 45 $ 125 Areca guppyana $ 30 $ 45 Areca ipot $ 45 Areca triandra $ 25 $ 30 $ 95 $ 125 Areca vestiaria $ 25 $ 30-35 $ 35-40 $ 55 $ 85-95 $ 125 Arecastrum romanzoffianum $ 125 Arenga australasica $ 20 $ 30 $ 35 $ 45-55 $ 85 $ 125 Arenga caudata $ 20 $ 30 $ 45 $ 55 $ 75 $ 100 Arenga engleri $ 20 $ 60 $ 35 $ 45 $ 85 $ 125 $ 200 $ 300+ Arenga hastata $ 25 www.junglemusic.net Page 1 of 22 Tree Citrus 25/+ Band$ 1 gal$ 2 gal$ 3/5 gal$ 7 gal$ 15 gal$ 20 gal$ Box$ Species Pot$ Pot$ gal$ Arenga hookeriana inquire Arenga micranthe 'Lhutan' $ 20 inquire Arenga pinnata $ 35 $ 50 $ 85 $ 125 Arenga sp. -

A Taxonomic Revision of Araceae Tribe Potheae (Pothos, Pothoidium and Pedicellarum) for Malesia, Australia and the Tropical Western Pacific
449 A taxonomic revision of Araceae tribe Potheae (Pothos, Pothoidium and Pedicellarum) for Malesia, Australia and the tropical Western Pacific P.C. Boyce and A. Hay Abstract Boyce, P.C. 1 and Hay, A. 2 (1Herbarium, Royal Botanic Gardens, Kew, Richmond, Surrey, TW9 3AE, U.K. and Department of Agricultural Botany, School of Plant Sciences, The University of Reading, Whiteknights, P.O. Box 221, Reading, RS6 6AS, U.K.; 2Royal Botanic Gardens, Mrs Macquarie’s Road, Sydney, NSW 2000, Australia) 2001. A taxonomic revision of Araceae tribe Potheae (Pothos, Pothoidium and Pedicellarum) for Malesia, Australia and the tropical Western Pacific. Telopea 9(3): 449–571. A regional revision of the three genera comprising tribe Potheae (Araceae: Pothoideae) is presented, largely as a precursor to the account for Flora Malesiana; 46 species are recognized (Pothos 44, Pothoidium 1, Pedicellarum 1) of which three Pothos (P. laurifolius, P. oliganthus and P. volans) are newly described, one (P. longus) is treated as insufficiently known and two (P. sanderianus, P. nitens) are treated as doubtful. Pothos latifolius L. is excluded from Araceae [= Piper sp.]. The following new synonymies are proposed: Pothos longipedunculatus Ridl. non Engl. = P. brevivaginatus; P. acuminatissimus = P. dolichophyllus; P. borneensis = P. insignis; P. scandens var. javanicus, P. macrophyllus and P. vrieseanus = P. junghuhnii; P. rumphii = P. tener; P. lorispathus = P. leptostachyus; P. kinabaluensis = P. longivaginatus; P. merrillii and P. ovatifolius var. simalurensis = P. ovatifolius; P. sumatranus, P. korthalsianus, P. inaequalis and P. jacobsonii = P. oxyphyllus. Relationships within Pothos and the taxonomic robustness of the satellite genera are discussed. Keys to the genera and species of Potheae and the subgenera and supergroups of Pothos for the region are provided.