Species Divers. 20(1): 67-71 (2015)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Mt.Iwaki Skyline International Challenge 2019 September 9Th
2019 青森岩木山國際單車挑戰日 Mt.Iwaki Skyline International Challenge 2019 September 9th Located at Mt Iwaki, a prominent mountain in Japan, this bike climb challenges all participants to bike their way up 15km of well maintained and symmetrical 69 hairpin turns. This route is only open during special events and races, take this opportunity and join us in climbing this spectacular route! EVENT LOCATION Tsugaru Iwaki Skyline Kuromori-56-2 Tokiwano, Hirosaki-shi, Aomori-ken 036-1345, Japan EVENT TIME 07:30 - 12:00 ROUTE DETAILS Mount Iwaki Parking Lot → Tsugaru Iwaki SkyLine 10 km hill climb | Base elevation 44m | Top elevation 1238m | Elevation difference 798m Max slope gradient 11.8% | Min slope gradient 4.5% | Average slope gradient 8.1% Mount Iwaki Skyline Arial Map Mount Iwaki Hill Climb Event Schedule Date Time Details 2019 SEPT 9 07:30 - 8:20 Report to check in area 8:20 Last call for check in; participants gather at the starting line 08:30 Challenge begins 08:30 - 11:00 Allotted challenge duration 11:00 Challenge officially closes 11:30 Ordered group descent starts Note 1 : If you finish the challenge climb early, you can take the cable car and enjoy a 30 - 40 minute hike up to a beautiful viewpoint on Tsugaru Fuji. 2 : If your team arranges your own caravan to accompany the ride, the caravan must reach the top of the mountain before 08:20. Caravans descent last, behind all participants. Challenge Rules 1. Participants must follow and complete the designated route. 2. This event is a challenge, not a race. -

HIRATA KOKUGAKU and the TSUGARU DISCIPLES by Gideon
SPIRITS AND IDENTITY IN NINETEENTH-CENTURY NORTHEASTERN JAPAN: HIRATA KOKUGAKU AND THE TSUGARU DISCIPLES by Gideon Fujiwara A THESIS SUBMITTED IN PARTIAL FULFILLMENT OF THE REQUIREMENTS FOR THE DEGREE OF DOCTOR OF PHILOSOPHY in The Faculty of Graduate Studies (Asian Studies) THE UNIVERSITY OF BRITISH COLUMBIA (Vancouver) April 2013 © Gideon Fujiwara, 2013 ABSTRACT While previous research on kokugaku , or nativism, has explained how intellectuals imagined the singular community of Japan, this study sheds light on how posthumous disciples of Hirata Atsutane based in Tsugaru juxtaposed two “countries”—their native Tsugaru and Imperial Japan—as they transitioned from early modern to modern society in the nineteenth century. This new perspective recognizes the multiplicity of community in “Japan,” which encompasses the domain, multiple levels of statehood, and “nation,” as uncovered in recent scholarship. My analysis accentuates the shared concerns of Atsutane and the Tsugaru nativists toward spirits and the spiritual realm, ethnographic studies of commoners, identification with the north, and religious thought and worship. I chronicle the formation of this scholarly community through their correspondence with the head academy in Edo (later Tokyo), and identify their autonomous character. Hirao Rosen conducted ethnography of Tsugaru and the “world” through visiting the northern island of Ezo in 1855, and observing Americans, Europeans, and Qing Chinese stationed there. I show how Rosen engaged in self-orientation and utilized Hirata nativist theory to locate Tsugaru within the spiritual landscape of Imperial Japan. Through poetry and prose, leader Tsuruya Ariyo identified Mount Iwaki as a sacred pillar of Tsugaru, and insisted one could experience “enjoyment” from this life and beyond death in the realm of spirits. -

Hachinohe Martial Arts Center About 15 Min
Access A Maeda Arena About 35 min. by car from Aomori Airport (New Aomori Prefecture Sports Park) About 25 min. by car from Shin-Aomori Station (JR Tohoku Shinkansen Line/Ou Line) B Michinokubank Dream Stadium About 30 min. by car from Aomori Airport (Aomori City Sports Complex) About 15 min. by car from Shin-Aomori Station (JR Tohoku Shinkansen Line/Ou Line) C Aomori Martial Arts Hall About 50 min. by car from Aomori Airport Hirosaki Athletic Park About 10 min. by car from Hirosaki Station (JR Ou Line/Konan Railway Konan Line) About 70 min. by car from Aomori Airport Mt. Iwakisan Synthesis Park About 40 min. by car from Hirosaki Station (JR Ou Line/Konan Railway Konan Line) D The right choice to train About 70 min. by car from Aomori Airport Iwaki Seishonen Sports Center About 45 min. by car from Hirosaki Station (JR Ou Line/Konan Railway Konan Line) AOMORI E About 70 min. by car from Aomori Airport Iwaki River Canoe Center About 30 min. by car from Hirosaki Station (JR Ou Line/Konan Railway Konan Line) About 55 min. by car from Misawa Airport Takamori Yama Sports Park About 30 min. by car from Shichinohe-Towada Station (JR Tohoku Shinkansen Line) F About 45 min. by car from Misawa Airport Towada City Wakaba Stadium About 25 min. by car from Shichinohe-Towada Station (JR Tohoku Shinkansen Line) About 40 min. by car from Misawa Airport Hachinohe Martial Arts Center About 15 min. by car from Hachinohe Station (JR Tohoku Shinkansen Line/Hachinohe Line/Aoimori Railway Line) G About 50 min. -

Japanese Landscapes: Where Land and Culture Merge
University of Kentucky UKnowledge Asian History History 1998 Japanese Landscapes: Where Land and Culture Merge Cotton Mather P. P. Karan University of Kentucky Shigeru Iijima Tokyo Institute of Ethnology, Japan Click here to let us know how access to this document benefits ou.y Thanks to the University of Kentucky Libraries and the University Press of Kentucky, this book is freely available to current faculty, students, and staff at the University of Kentucky. Find other University of Kentucky Books at uknowledge.uky.edu/upk. For more information, please contact UKnowledge at [email protected]. Recommended Citation Mather, Cotton; Karan, P. P.; and Iijima, Shigeru, "Japanese Landscapes: Where Land and Culture Merge" (1998). Asian History. 2. https://uknowledge.uky.edu/upk_asian_history/2 JAPANESE LANDSCAPES JAPANESE LANDSCAPES Where Land & Culture Merge by Cotton Mather, EE Karan, and Shigeru Iijima THE UNIVERSITY PRESS OF KENTUCKY Copyright © 1998 by The University Press of Kentucky Field research in Japan and preparation ofillustrations for this Scholarly publisher for the Commonwealth, serving Bellarmine College, book were supported by the Berea College, Centre College of Kentucky, Eastern Kentucky University, Geographical Society, Mesilla, The Filson Club Historical Society, Georgetown College, Kentucky Historical New Mexico. Society, Kentucky State University, Morehead State University, Murray State University, Northern Kentucky University, Transylvania University, University Publication ofthis volume was of Kentucky, University of Louisville, and Western Kentucky University. made possible in part by a grant All rights reserved from the National Endowment for the Humanities. Editorial and Sales offices: The University Press of Kentucky 663 South Limestone Street, Lexington, Kentucky 40508-4008 02 01 00 99 98 5 4 3 2 I LIBRARY OF CONGRESS CATALOGING-IN-PUBLICATION DATA Mather, Cotton, 1918- Japanese landscapes: where land & culture merge / by Cotton Mather, ~~ Karan, and Shigeru Iijima. -

Historical Fish Specimens Collected from the Tohoku District by the Saito Ho-On Kai Museum of Natural History
Bull. Natl. Mus. Nat. Sci., Ser. A, 35(1), pp. 9–54, March 22, 2009 Historical Fish Specimens Collected from the Tohoku District by the Saito Ho-on Kai Museum of Natural History Keiichi Matsuura1, Gento Shinohara2 and Masanori Nakae1 1 Collection Center, National Museum of Nature and Science, 3–23–1 Hyakunin-cho, Shinjuku-ku, Tokyo, 169–0073 Japan E-mail: [email protected]; [email protected] 2 Department of Zoology, National Museum of Nature and Science, 3–23–1 Hyakunin-cho, Shinjuku-ku, Tokyo, 169–0073 Japan E-mail: [email protected] Abstract The fish collection of the Saito Ho-on Kai Museum of Natural History was transferred to the National Museum of Nature and Science, Tokyo in February 2006. Ninety percent of the fish collection contains specimens collected from the Tohoku District during the period from 1930 to 1933 when natural environments of Japan were in good condition for various groups of fishes. The fish specimens from the Tohoku District were classified into 361 species/subspecies of 273 genera belonging to 131 families of 31 orders. A list of the species is shown with remarks on distribution. Key words: Fish specimens, Saito Ho-on Kai Museum, Tohoku District, inventory. stead of natural sicence. The museum has tried to Introduction keep its activity at the level before the war, but it The Saito Ho-on Kai Museum was established failed to do so because of financial difficulties. In in November 1933 in Sendai City, Miyagi Pre- 2005, the Saito Ho-on Kai Museum of Natural fecture, Japan. -
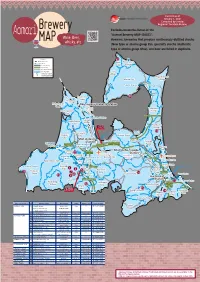
Wine, Beer, Whisky, Etc
Current as of January 1, 2021 Compiled by Sendai Regional Taxation Bureau Brewery Excludes breweries listed on the Aomori Wine, Beer, "Aomori Brewery MAP (SAKE)". MAP whisky, etc. However, breweries that produce continuously distilled shochu (New type or shochu group Ko), specialty shochu (Authentic type or shochu group Otsu), and beer are listed in duplicate. Legend ㉑ 279 Shinkansen Kazamaura JR Line Oma Private Railway Town Village Expressway Major National Highway City Boundary Sai Shinkansen Station JR and Private Railway Stations Village Higashidoori Mutsu City Village Ominato Station Shimokita Station 279 Ominato Line Imabetsu 338 ⑳ Town Sotogahama Town Nakadomari Okutsugaru-Imabetsu Station Town 338 Sotogahama Yokohama Goshogawara TsugaruTown Line Town City Kanita Station Yomogita Nakadomari Village Town Hokkaido Shinkansen Rokkasho 339 280 ①② Village 4 Noheji ⑲ Goshogawara Aomori Port Station Tsugaru City Hiranai Town City Shin-Aomori Aomori Station Noheji Town Tohoku Ajigasawa Station 101 Goshogawara Town Station Aoimori Railway394 Line Station Ou Misawa Main 103 4 Line Shichinohe-Towada City Tsuruta 338 Town ⑰ Aomori Airport Station Itayanagi Shichinohe Misawa Airport Itayanagi Town Namioka Station Aomori City Town Tohoku Shinkansen Station 394 Misawa Station Ajigasawa Town Fujisaki Kuroishi Kawabe Station Town Station 394 Oirase Town Fukaura Station Hirosaki Inakadate Hachinohe Port City ④ VillageKonan ③ Railway Kuroishi 102 Rokunohe 45 Gono Line Fukaura ⑤ City Hon- ⑯ ⑦ Hiraka 4 Town Town Towada City ⑱ Hachinohe Same Station ⑧ Station Station ⑪ ⑨ 454 Hachinohe Hirosaki Station Station ⑬ Mutsu-Minato Station Nishimeya Hirakawa Hachinohe Line Village Gonohe Town Owani⑫ City 454 ⑥⑩ Town ⑮ 7 282 Shingo Village ⑭ Hashikami Nanbu Hachinohe Town Town City Sannohe Town Name of City, Town, and Village No. -

Mutsu ONE Trail
Imabetsu Town Tsugaruhamana Station Imabetsu Town Tsugaruhamana Station Sotogahama Town Top Mast Aomori City WA-RASSE A full 250-km walking Mutsu ONE Trail trip around Mutsu Bay ̶8 Model Routes to Enjoy the Mutsu Bay̶ Sotogahama Town Tappizaki Sotogahama Town Top Mast Aomori City ASPAM Aomori City Asamushi Onsen Designated as Tsugaru Quasi-National Park, this area Takanozaki in Imabetsu is a stunning, scenic spot with Tamamatsu Beach in Yomogita is widely known for its You can walk through Aoiumi Park in Aomori while This leaflet introduces the eight model routes to enjoy the “Mutsu allows you to walk all the way while enjoying the sea mysterious-looking and oddly-shaped rocks jutting out everywhere. “crystal-clear water.” Between Yomogita and Aburakawa enjoying a gentle sea breeze and a view of the Mutsu Mutsu ONE Trail The one-kilometer row of black pines in Tairadate, Sotogahama, is ONE Trail.” There are a myriad of ways to enjoy it, depending on breeze nearby. You can see Hokkaido to the north, in Aomori, the road runs closely along the sea, allowing Bay. The sunset from Asamushi Beach will fascinate called “Matsumae Kaido” because the Matsumae clan used to you to walk while viewing the Shimokita Peninsula to the Suggested Travel Itineraries when and where you visit. You can start from any point according Tappizaki to the west, and the Shimokita Peninsula to travel along this road during the Edo period under the Tokugawa visitors, because it looks as if the sun is being drawn into north, Asamushi to the west on other side of the coast, the east. -

By Municipality) (As of March 31, 2020)
The fiber optic broadband service coverage rate in Japan as of March 2020 (by municipality) (As of March 31, 2020) Municipal Coverage rate of fiber optic Prefecture Municipality broadband service code for households (%) 11011 Hokkaido Chuo Ward, Sapporo City 100.00 11029 Hokkaido Kita Ward, Sapporo City 100.00 11037 Hokkaido Higashi Ward, Sapporo City 100.00 11045 Hokkaido Shiraishi Ward, Sapporo City 100.00 11053 Hokkaido Toyohira Ward, Sapporo City 100.00 11061 Hokkaido Minami Ward, Sapporo City 99.94 11070 Hokkaido Nishi Ward, Sapporo City 100.00 11088 Hokkaido Atsubetsu Ward, Sapporo City 100.00 11096 Hokkaido Teine Ward, Sapporo City 100.00 11100 Hokkaido Kiyota Ward, Sapporo City 100.00 12025 Hokkaido Hakodate City 99.62 12033 Hokkaido Otaru City 100.00 12041 Hokkaido Asahikawa City 99.96 12050 Hokkaido Muroran City 100.00 12068 Hokkaido Kushiro City 99.31 12076 Hokkaido Obihiro City 99.47 12084 Hokkaido Kitami City 98.84 12092 Hokkaido Yubari City 90.24 12106 Hokkaido Iwamizawa City 93.24 12114 Hokkaido Abashiri City 97.29 12122 Hokkaido Rumoi City 97.57 12131 Hokkaido Tomakomai City 100.00 12149 Hokkaido Wakkanai City 99.99 12157 Hokkaido Bibai City 97.86 12165 Hokkaido Ashibetsu City 91.41 12173 Hokkaido Ebetsu City 100.00 12181 Hokkaido Akabira City 97.97 12190 Hokkaido Monbetsu City 94.60 12203 Hokkaido Shibetsu City 90.22 12211 Hokkaido Nayoro City 95.76 12220 Hokkaido Mikasa City 97.08 12238 Hokkaido Nemuro City 100.00 12246 Hokkaido Chitose City 99.32 12254 Hokkaido Takikawa City 100.00 12262 Hokkaido Sunagawa City 99.13 -

Aomori Cycling
AOMORI CYCLING Up close and personal with the incredible nature and culture of Aomori Aomori Cycling Up close and personal with the incredible nature and culture of Aomori Surrounded by ocean on all sides and blessed by incredible nature such as Shirakami-Sanchi and Lake Towada, each region in Aomori Prefecture enjoys its own unique history, culture and delicacies. Cycling Aomori is the perfect way to discover the intricacies of these individual regions. If you are ready, then jump on your bike and let's head off to explore the beauty of Aomori! 9 1 Course 1 NATSUDOMARI 11 2 Course 2 TOWADA・OIRASE 13 3 Course 3 HIROSAKI 15 4 Course 4 NISHIKAIGAN 17 5 Course 5 OKU-TSUGARU AOMORI 19 6 Course 6 SHIMOKITA 21 7 Course 7 LAKE OGAWARA CYCLING 23 Course 8 HACHINOHE Up close and personal with the incredible nature and culture of Aomori 8 25 33 27 35 29 37 31 7 8 03 02 04 05 06 01/The masses of blue net stored at the port are shellfish baskets, used for cultivating scallops. It's like riding through a maze. 02/Old fishing vessels of all shapes and sizes are dotted around the Natsudomari Peninsula. 03/Showing off the size of the scallops at "Hotate Hiroba," which has a giant scallop as its signboard. You can learn about scallop cultivation on the second floor.04 /At Yogoshiyama Forest Park you can see over 3,000 varieties of succulents. 05/Two men and a cat taking a break from cycling beside the tetrapod seawall. -

Distribution of the Japanese Yellow Bunting Emberiza Sulphurata on Shimokita and Tsugaru Peninsulas, Aomori Prefecture 総合政策 第○巻第○号(201●) 総合政策 第○巻第○号(201●)
View metadata, citation and similar papers at core.ac.uk brought to you by CORE 短報 Distribution総合政策 第 of the Japanese 21 巻(2020)pp.109-114 yellow bunting Emberiza sulphurata on Shimokita andJournal Tsugaru of Peninsulas,Policy Studies Aomori Prefecture B5版 新様式 総合政策 第○巻第○号(201●) pp.1-20 Journal of Policy Studies Distribution of the Japanese yellow bunting Emberiza sulphurata on Shimokita and Tsugaru Peninsulas, Aomori Prefecture ✻ Yoshio Kaneko Abstract The Japanese yellow bunting Emberiza sulphurata breeds only in Japan and is distributed mainly in northern Japan. I made observations on the species on Shimokita and Tsugaru Peninsulas which are the northernmost parts of mainland Honshu. The species was frequently found in both peninsulas and occurred in areas lower than 60 m in altitude. According to the IUCN Redlist, the species breeds from 600 m to 1,500 m. Since it can be found in the areas lower than 600 m on Shimokita, Tsugaru and elsewhere, IUCN’s description needs to be rectified. Considering the occurrence in a wide range of altitudes, the population of the Japanese yellow bunting may be larger than once believed. K ey words northern Japan, altitudinal distribution, habitat, IUCN Redlist Introduction The Japanese yellow bunting Emberiza sulphurata is a small passerine bird species belonging to the family Emberizidae (Fig. 1). The species is known to breed only in Japan. It is distributed and breeds mainly in northern Japan. There is no breeding record from Hokkaido. It used to be thought to breed in limited areas such as Mt. Fuji. However, it has recently become known that the species is fairly common in northern Japan, especially in the Fig. -

A Prosperous Future Starts Here
A prosperous future starts here 100% of this paper was made using recycled paper 2018.4 (involved in railway construction) Table of Lines Constructed by the JRTT Contents Tsukuba Tokyo Area Lines Constructed by JRTT… ……………………… 2 Sassho Line Tsukuba Express Line Asahikawa Uchijuku JRTT Main Railway Construction Projects……4 Musashi-Ranzan Signal Station Saitama Railway Line Maruyama Hokkaido Shinkansen Saitama New Urban Musashino Line Tobu Tojo Line Urawa-Misono Kita-Koshigaya (between Shin-Hakodate-Hokuto Transit Ina Line Omiya Nemuro Line Shinrin-Koen and Sapporo) ■ Comprehensive Technical Capacity for Railway Sapporo Construction/Research and Plans for Railway Tobu Isesaki Line Narita SKY ACCESS Line Construction… ………………………………………………6 Hatogaya (Narita Rapid Rail Acess Line) Shiki Shin-Matsudo Hokuso Railway Hokuso Line ■ Railway Construction Process… …………………………7 Takenotsuka Tobu Tojo Line Shin-Kamagaya Komuro Shin-Hakodatehokuto Seibu Wako-shi Akabane Ikebukuro Line Imba Nihon-Idai Sekisho Line Higashi-Matsudo Narita Airport Hakodate …… Kotake-Mukaihara Toyo Rapid Construction of Projected Shinkansen Lines 8 Shakujii-Koen Keisei-Takasago Hokkaido Shinkansen Aoto Nerima- Railway Line Nerima Takanodai Ikebukuro Keisei Main Line (between Shin-Aomori and Shin-Hakodate-Hokuto) Hikifune Toyo- Tsugaru-Kaikyo Line Seibu Yurakucho Line Tobu Katsutadai ■ Kyushu Shinkansen… ………………………………………9 Tachikawa Oshiage Ueno Isesaki Line Keio Line Akihabara Nishi-Funabashi Shinjuku … ………………………………… Odakyu Odawara Line Sasazuka ■ Hokuriku Shinkansen 10 Yoyogi-Uehara -

TSUGARU AREA AREA CYCLING COURSE MAP CYCLING COURSE MAP Feel the Wind of Aomori
TSUGARU TSUGARU AREA AREA CYCLING COURSE MAP CYCLING COURSE MAP Feel the wind of Aomori. Cycling Journey to discover the great view, food and culture. 1 TSUGARU AREA CYCLING COURSE MAP 2 TSUGARU AREA CYCLING COURSE MAP 1 INTRODUCTION 5 COURSE 1 ALL AROUND TSUGARU PENINSULA 7 COURSE 2 MOUNTAIN CHALLENGE 9 COURSE 3 Mt.MANOGAMI 11 COURSE 4 APPLE ORCHARD COURSE 5 AROUND LAKE JUSANKO 13 $0634& Imabetsu-town 15 COURSE 6 TSURU NO MAI BRIDGE Sotogahama-town 17 COURSE 7 IWAKI RIVER Goshogawara-city $0634& 19 COURSE 8 AROUND OKUTSUGARU Lake COURSE 9 TSUGARU STRAIT Jusanko 21 $0634& COURSE 10 SEA OF JAPAN Yomogita-village 23 Nakadomari-town COURSE 11 BLUE POND 25 $0634& COURSE 12 WORLD HERITAGE SHIRAKAMI 27 $0634& Goshogawara-city 29 MAIN SPOT & EVENT AROUND TSUGARU Tsugaru-city 33 SPECIAL SERVICE FOR CYCLIST $0634& $0634& Tsuruta-town Aomori-city 35 TRAFFIC RULES & MANNERS IN ORDER TO ENJOY AOMORI $0634& 36 $0634& $0634& 37 INFORMATION Ajigasawa-town Mt.Iwaki Fukaura-town $0634& Hirosaki-city Shirakami- $0634& Sanchi Nishimeya-village 1 2 3 TSUGARU AREA CYCLING COURSE 5 Tappi-zaki Takano-saki Cape Cape e Minmaya Bay in L i r ALL AROUND a Chokandai m o Observatory Strait Tairadate d u s Imabetsu-town t a Tsugaru Line T Rest Area TSUGARU PENINSULA 339 Kodomari Sotogahama-town "PONTOMARI" 280 Goshogawara-city 4 6 Full-loaded Coastline Ride Lake Jusanko Sea of Japan Yomogita-village Hokkaido Shinkansen Takayama Inari Nakadomari-town Iwaki River Tsugaru Railway Line Shrine Marche 200km challenge ride going ”YOMOGITA” 12 around Tsugaru Peninsula Monument of ASUNARO-Line This is a challenging course that you pedal for 200km and go around Tsugaru Peninsula.