Fig. 1-5. Trees Or Shrubs, Evergreen (Mal. Spp.)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Alphabetical Lists of the Vascular Plant Families with Their Phylogenetic
Colligo 2 (1) : 3-10 BOTANIQUE Alphabetical lists of the vascular plant families with their phylogenetic classification numbers Listes alphabétiques des familles de plantes vasculaires avec leurs numéros de classement phylogénétique FRÉDÉRIC DANET* *Mairie de Lyon, Espaces verts, Jardin botanique, Herbier, 69205 Lyon cedex 01, France - [email protected] Citation : Danet F., 2019. Alphabetical lists of the vascular plant families with their phylogenetic classification numbers. Colligo, 2(1) : 3- 10. https://perma.cc/2WFD-A2A7 KEY-WORDS Angiosperms family arrangement Summary: This paper provides, for herbarium cura- Gymnosperms Classification tors, the alphabetical lists of the recognized families Pteridophytes APG system in pteridophytes, gymnosperms and angiosperms Ferns PPG system with their phylogenetic classification numbers. Lycophytes phylogeny Herbarium MOTS-CLÉS Angiospermes rangement des familles Résumé : Cet article produit, pour les conservateurs Gymnospermes Classification d’herbier, les listes alphabétiques des familles recon- Ptéridophytes système APG nues pour les ptéridophytes, les gymnospermes et Fougères système PPG les angiospermes avec leurs numéros de classement Lycophytes phylogénie phylogénétique. Herbier Introduction These alphabetical lists have been established for the systems of A.-L de Jussieu, A.-P. de Can- The organization of herbarium collections con- dolle, Bentham & Hooker, etc. that are still used sists in arranging the specimens logically to in the management of historical herbaria find and reclassify them easily in the appro- whose original classification is voluntarily pre- priate storage units. In the vascular plant col- served. lections, commonly used methods are systema- Recent classification systems based on molecu- tic classification, alphabetical classification, or lar phylogenies have developed, and herbaria combinations of both. -

Evolutionary History of Floral Key Innovations in Angiosperms Elisabeth Reyes
Evolutionary history of floral key innovations in angiosperms Elisabeth Reyes To cite this version: Elisabeth Reyes. Evolutionary history of floral key innovations in angiosperms. Botanics. Université Paris Saclay (COmUE), 2016. English. NNT : 2016SACLS489. tel-01443353 HAL Id: tel-01443353 https://tel.archives-ouvertes.fr/tel-01443353 Submitted on 23 Jan 2017 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. NNT : 2016SACLS489 THESE DE DOCTORAT DE L’UNIVERSITE PARIS-SACLAY, préparée à l’Université Paris-Sud ÉCOLE DOCTORALE N° 567 Sciences du Végétal : du Gène à l’Ecosystème Spécialité de Doctorat : Biologie Par Mme Elisabeth Reyes Evolutionary history of floral key innovations in angiosperms Thèse présentée et soutenue à Orsay, le 13 décembre 2016 : Composition du Jury : M. Ronse de Craene, Louis Directeur de recherche aux Jardins Rapporteur Botaniques Royaux d’Édimbourg M. Forest, Félix Directeur de recherche aux Jardins Rapporteur Botaniques Royaux de Kew Mme. Damerval, Catherine Directrice de recherche au Moulon Président du jury M. Lowry, Porter Curateur en chef aux Jardins Examinateur Botaniques du Missouri M. Haevermans, Thomas Maître de conférences au MNHN Examinateur Mme. Nadot, Sophie Professeur à l’Université Paris-Sud Directeur de thèse M. -

Phylogenetic Relationships in the Order Ericales S.L.: Analyses of Molecular Data from Five Genes from the Plastid and Mitochondrial Genomes1
American Journal of Botany 89(4): 677±687. 2002. PHYLOGENETIC RELATIONSHIPS IN THE ORDER ERICALES S.L.: ANALYSES OF MOLECULAR DATA FROM FIVE GENES FROM THE PLASTID AND MITOCHONDRIAL GENOMES1 ARNE A. ANDERBERG,2,5 CATARINA RYDIN,3 AND MARI KAÈ LLERSJOÈ 4 2Department of Phanerogamic Botany, Swedish Museum of Natural History, P.O. Box 50007, SE-104 05 Stockholm, Sweden; 3Department of Systematic Botany, University of Stockholm, SE-106 91 Stockholm, Sweden; and 4Laboratory for Molecular Systematics, Swedish Museum of Natural History, P.O. Box 50007, SE-104 05 Stockholm, Sweden Phylogenetic interrelationships in the enlarged order Ericales were investigated by jackknife analysis of a combination of DNA sequences from the plastid genes rbcL, ndhF, atpB, and the mitochondrial genes atp1 and matR. Several well-supported groups were identi®ed, but neither a combination of all gene sequences nor any one alone fully resolved the relationships between all major clades in Ericales. All investigated families except Theaceae were found to be monophyletic. Four families, Marcgraviaceae, Balsaminaceae, Pellicieraceae, and Tetrameristaceae form a monophyletic group that is the sister of the remaining families. On the next higher level, Fouquieriaceae and Polemoniaceae form a clade that is sister to the majority of families that form a group with eight supported clades between which the interrelationships are unresolved: Theaceae-Ternstroemioideae with Ficalhoa, Sladenia, and Pentaphylacaceae; Theaceae-Theoideae; Ebenaceae and Lissocarpaceae; Symplocaceae; Maesaceae, Theophrastaceae, Primulaceae, and Myrsinaceae; Styr- acaceae and Diapensiaceae; Lecythidaceae and Sapotaceae; Actinidiaceae, Roridulaceae, Sarraceniaceae, Clethraceae, Cyrillaceae, and Ericaceae. Key words: atpB; atp1; cladistics; DNA; Ericales; jackknife; matR; ndhF; phylogeny; rbcL. Understanding of phylogenetic relationships among angio- was available for them at the time, viz. -

Common Name Scientific Name Type Plant Family Native
Common name Scientific name Type Plant family Native region Location: Africa Rainforest Dragon Root Smilacina racemosa Herbaceous Liliaceae Oregon Native Fairy Wings Epimedium sp. Herbaceous Berberidaceae Garden Origin Golden Hakone Grass Hakonechloa macra 'Aureola' Herbaceous Poaceae Japan Heartleaf Bergenia Bergenia cordifolia Herbaceous Saxifragaceae N. Central Asia Inside Out Flower Vancouveria hexandra Herbaceous Berberidaceae Oregon Native Japanese Butterbur Petasites japonicus Herbaceous Asteraceae Japan Japanese Pachysandra Pachysandra terminalis Herbaceous Buxaceae Japan Lenten Rose Helleborus orientalis Herbaceous Ranunculaceae Greece, Asia Minor Sweet Woodruff Galium odoratum Herbaceous Rubiaceae Europe, N. Africa, W. Asia Sword Fern Polystichum munitum Herbaceous Dryopteridaceae Oregon Native David's Viburnum Viburnum davidii Shrub Caprifoliaceae Western China Evergreen Huckleberry Vaccinium ovatum Shrub Ericaceae Oregon Native Fragrant Honeysuckle Lonicera fragrantissima Shrub Caprifoliaceae Eastern China Glossy Abelia Abelia x grandiflora Shrub Caprifoliaceae Garden Origin Heavenly Bamboo Nandina domestica Shrub Berberidaceae Eastern Asia Himalayan Honeysuckle Leycesteria formosa Shrub Caprifoliaceae Himalaya, S.W. China Japanese Aralia Fatsia japonica Shrub Araliaceae Japan, Taiwan Japanese Aucuba Aucuba japonica Shrub Cornaceae Japan Kiwi Vine Actinidia chinensis Shrub Actinidiaceae China Laurustinus Viburnum tinus Shrub Caprifoliaceae Mediterranean Mexican Orange Choisya ternata Shrub Rutaceae Mexico Palmate Bamboo Sasa -

Clethra, Or Summersweet, Attracts Butterflies and Numerous Pollinators and Is a Great Alternative to the Invasive Butterfly Bush
Share this: Clethra, or Summersweet, attracts butterflies and numerous pollinators and is a great alternative to the invasive butterfly bush. Its bright green leaves are followed by fragrant spires of flowers in mid to late summer. Here at the nursery, they are about to pop into bloom any day now. We carry the following selections. All Clethra prefer a moist to wet site, sun to light shade. Clethra alnifolia: Grows 4' or more, white blossoms. One of the easiest butterfly shrubs you can have. This is the 'straight species' that occurs naturally in low areas around woodlands and wetlands. Clethra alnifolia 'Ruby Spice': This selection bears deep rose-colored, non-fading fragrant flowers. 6'-8' high. Clethra alnifolia 'Hummingbird': A dwarf selection that grows 2-1/2 to 3-1/2 feet tall. Clethra alnifolia 'Sixteen Candles'" Lustrous dark green foliage, 4" to 6" racemes are held upright like birthday candles (pictured on far left). The shrub reaches 3'-5'. SUMMER BLOOMING PERENNIALS If you have not visited the nursery yet, now is the time. Summer is when native plants out-do themselves with color. Combined with the many insects and birds they support, the show is non-stop. Visit our demonstration gardens and view the selection of over 300 native species we offer for sale. Many of our plants are seed grown to enhance genetic diversity in the ecosystem. What People Are Saying About Classes at Edge of the Woods "Thanks so much! I had a great time and learned a lot"! "Everyone was so friendly and helpful. I'm glad I came." "I am so glad I came, I never visited before and will definitely be back." Edge of the Woods Native Plant Nursery is a women-owned, independent business. -

Japanese Clethra: a Hidden Gem
Japanese Clethra: A Hidden Gem Richard Schulhof CHULHOF t the edge of the Arnold Arboretum’s S Central Woods, far from most visitors, ICHARD Agrows an exceptional specimen of Jap- R anese clethra (Clethra barbinervis, accession 13087). I first became enamored with this spe- cies as a student at Longwood Gardens, where I admired its elegant form as part of the backdrop to the famed Flower Garden Walk. While I later saw several very fine examples of Japanese clethra in the great gardens of the Delaware Valley, none approached the singular beauty and character of the Arnold Arboretum’s specimen. Like many plants in the Arboretum’s collec- tions, this accession comes with an impressive pedigree, tracing back to Japan in 1886. In that Japanese clethra’s leaves are deep green year, William Penn Brooks, a Massachusetts through summer. Autumn color tends to be native and valedictorian of the state agricultural unreliable. I have seen outstanding tints of red college class of 1875, sent seeds of several species and burgundy on specimens in both south- to the Arnold Arboretum. Brooks, then a teacher eastern Pennsylvania and here at the Arnold and administrator at Sapporo Agricultural Arboretum, but in other years have noted little School, found time to survey the surrounding color change before the leaves fall away incon- countryside of Hokkaido for interesting plants, spicuously. Japanese clethra’s white flowers are several of which came to enrich the Arboretum, similar to those of summersweet, but they are including katsura tree (Cercidiphyllum japoni- borne in 4 to 6 inch long panicled racemes that cum) and hardy kiwi (Actinidia arguta). -
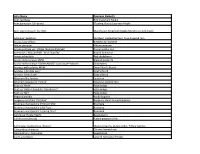
Latin Name Common Name(S) Acer Capillipes Red Snakebark Maple Acer Palmatum 'Uki-Gumo' Floating Clouds Japanese Maple
Latin Name Common Name(s) Acer capillipes Red Snakebark Maple Acer palmatum 'Uki-gumo' Floating Clouds Japanese Maple Acer tegmentosum 'Joe Witt' Manchurian Stripebark Maple,Manchu striped maple Adiantum pedatum Northern maidenhair fern, Five-fingered fern Aesculus parviflora Bottlebrush buckeye Allium cernuum Allium cernuum Amsonia ciliata var. filifolia 'Georgia Pancake' Creeping Blue Star Anemone x 'MACANE001' Wild SwanTM Hybrid Anemone Aronia arbutifolia Red chokeberry Aronia melanocarpa MVW Black chokeberry Aronia melanocarpa 'UCONNAM165' Low Scape® Mound Chokeberry Aruncus aethusifolius MVW Dwarf Goat's Beard Aruncus 'Chantilly Lace' Goat's Beard Aruncus 'Misty Lace' Goat's Beard Athyrium filix-femina Lady fern Athyrium niponicum 'Pictum' Japanese painted fern Baptisia 'Anne' False Indigo Baptisia 'Dutch Chocolate' Decadence® False Indigo Baptisia 'Nell' False Indigo Begonia grandis Hardy begonia Bergenia cordifolia 'Rotblum' Bergenia, Heart-leaved bergenia Brunnera macrophylla 'Emerald Mist' Brunnera Brunnera macrophylla 'Jack Frost' Brunnera Brunnera macrophylla 'Looking Glass' Brunnera Callicarpa 'Purple Pearls' Beautyberry Callirhoe involucrata Purple poppy mallow Callitropsis nootkatensis 'Glauca' Nootka cypress, Alaska-cedar, Yellow cypress Calycanthus chinensis Chinese Sweetshrub Calycanthus x 'Aphrodite' Sweetshrub Carex amphibola Eastern Narrowleaf Sedge Carex appalachica Appalachian sedge Carex cherokeensis Cherokee sedge Carex pensylvanica Pennsylvania sedge Carex siderosticha 'Banana Boat' Creeping broadleaf sedge Cephalotaxus -

Checklist of the Washington Baltimore Area
Annotated Checklist of the Vascular Plants of the Washington - Baltimore Area Part I Ferns, Fern Allies, Gymnosperms, and Dicotyledons by Stanwyn G. Shetler and Sylvia Stone Orli Department of Botany National Museum of Natural History 2000 Department of Botany, National Museum of Natural History Smithsonian Institution, Washington, DC 20560-0166 ii iii PREFACE The better part of a century has elapsed since A. S. Hitchcock and Paul C. Standley published their succinct manual in 1919 for the identification of the vascular flora in the Washington, DC, area. A comparable new manual has long been needed. As with their work, such a manual should be produced through a collaborative effort of the region’s botanists and other experts. The Annotated Checklist is offered as a first step, in the hope that it will spark and facilitate that effort. In preparing this checklist, Shetler has been responsible for the taxonomy and nomenclature and Orli for the database. We have chosen to distribute the first part in preliminary form, so that it can be used, criticized, and revised while it is current and the second part (Monocotyledons) is still in progress. Additions, corrections, and comments are welcome. We hope that our checklist will stimulate a new wave of fieldwork to check on the current status of the local flora relative to what is reported here. When Part II is finished, the two parts will be combined into a single publication. We also maintain a Web site for the Flora of the Washington-Baltimore Area, and the database can be searched there (http://www.nmnh.si.edu/botany/projects/dcflora). -

Wardani Et Al
Floribunda 4(5) 2012 107 ENDEMIC PLANTS OF MT. RINJANI: AN OUTLOOK TO THE CONSERVATION STRATEGY W Wardani, A Hidayat, EF Tihurua, A Kartonegoro, LD Sulistyaningsih, ES Kuncari & EB Walujo “Herbarium Bogoriense”, Bidang Botani, Pusat Penelitian Biologi, Cibinong Science Center – LIPI, Jl. Raya Jakarta-Bogor Km. 46, Cibinong, Bogor, 16911. [email protected] (corresponding author) W Wardani, A Hidayat, EF Tihurua, A Kartonegoro, LD Sulistyaningsih, ES Kuncari & EB Walujo. 2012. Tumbuhan Endemik di Gunung Rinjani: Tinjauan Terhadap Strategi Konservasi. Floribunda 4(5): 107–112. — Gunung Rinjani diyakini merupakan sumber utama keanekaragaman tumbuhan di Pulau Lombok. Hal ini disebabkan pesatnya perubahan fungsi lahan di dataran yang lebih rendah di pulau tersebut, terutama untuk kebutuhan pertanian. Walaupun jenis-jenis endemik diperkirakan kurang dari 50 % dari total jumlah jenis tumbuhan lokal, kelompok ini sangat penting untuk konservasi. Hal ini disebabkan kurangnya informasi yang telah diketahui terhadap jenis-jenis tersebut sedangkan pada saat yang sama habitatnya terancam oleh aktifitas pariwisata, terutama pada jalur yang menuju ke kaldera. Strategi konservasi yang memastikan ke- lestarian jenis-jenis ini perlu segera ditetapkan. Dalam tulisan ini dipaparkan kehadiran jenis-jenis endemik di Pulau Lombok berdasarkan rekaman spesimen herbarium dan berbagai pustaka, berikut usulan-usulan un- tuk disertakan dalam strategi konservasinya. Kata kunci: Endemik, Pulau Lombok, Gunung Rinjani, strategi konservasi. W Wardani, A Hidayat, EF Tihurua, A Kartonegoro, LD Sulistyaningsih, ES Kuncari & EB Walujo. 2012. Endemic Plants of Mt. Rinjani: An Outlook to the Conservation Strategy. Floribunda 4(5): 107–112. — Mount Rinjani, lies in the island of Lombok, is believed as the major source of plant diversity in the island today. -

Salvia Huastecana (Lamiaceae), a New Species from San Luis Potosí, Mexico
Phytotaxa 433 (1): 001–008 ISSN 1179-3155 (print edition) https://www.mapress.com/j/pt/ PHYTOTAXA Copyright © 2020 Magnolia Press Article ISSN 1179-3163 (online edition) https://doi.org/10.11646/phytotaxa.433.1.1 Salvia huastecana (Lamiaceae), a new species from San Luis Potosí, Mexico BRENDA Y. BEDOLLA-GARCÍA1, SERGIO ZAMUDIO2 & HUGO A. CASTILLO-GÓMEZ3 1Instituto de Ecología, A. C., Centro Regional del Bajío, Red de Diversidad Biológica del Occidente Mexicano. Apdo. Postal 386, 61600 Pátzcuaro, Michoacán, Mexico; [email protected] 2Apartado Postal 392, 61600 Pátzcuaro, Michoacán, Mexico; [email protected] 3Universidad Autónoma de Querétaro, Facultad de Ciencias, herbario QMEX, Juriquilla, Querétaro, Mexico; [email protected] Abstract Salvia huastecana is described and illustrated as a new species. It is known from the municipality of Xilitla, San Luis Potosí, Mexico. This taxon belongs to Salvia sect. Angulatae, is similar to S. albiflora and S. roscida but it can be distinguished by a higher number of flowers per verticillaster (9−12), longer caducous bracts with long caudate apex, hirsute calyx with erect trichomes and white corolla, sometimes with the upper lip and margins of the lateral lobes of the lower lip light blue stained. Photographs of living plants and the illustration of the new species are provided. Resumen Se describe e ilustra como especie nueva a Salvia huastecana, conocida del municipio de Xilitla, San Luis Potosí, México. Este taxon pertenece a Salvia secc. Angulatae, es similar a Salvia albiflora y S. roscida, pero se distingue por presentar mayor número de flores por verticilastro (9−12), brácteas caedizas, más largas y con el ápice largamente caudado, cáliz hirsuto con tricomas erectos y corola blanca, en ocasiones con el labio superior y el margen de los lóbulos laterales del labio inferior teñidos tenuemente de azul. -

Chart of Bee Forage Plants Suitable for Growing in East Texas TREES
Chart of Bee Forage Plants Suitable for Growing in East Texas TREES COMMON NAME FAMILY GENUS Red Maple Acerancea Acer Rubrum Dogwood Cornaceae Cornus Florida Vitex Verbenacea Vitex Agnus-Castus Willows Salicaceae Salix Magnolia Magnoliaceae Magnolia Crabapple Rosaceae Malus Ornamental Pears Rosaceae Pyrus Calleryana Ornamental Wild Plums Rosaceae Prunus Ornamental Cherry Rosaceae Prunus Desert Willow Bignoniaceae Chilopsis Redbud Fabaceae Cercis Ninebark Rosaceae Physocarpus Sumac Anacardiaceae Rhus Aromatica Black Locust Fabaceae Robinis Pseudoacacia Tulip Poplar Magnoliaceae Liriodendron Tulipifera Mesquite Fabaceae Prosopsis Glandulosa All Fruit and Citrus Trees SHRUBS COMMON NAME FAMILY GENUS Abelia Caprifoliaceae Abelia Azalea Ericaceae Azalea or Rhododenron Butterfly Bush Loganiaceae Buddleia Daviddi Barberry Berberidaceae Berberis Barrel Cactus Cactaceae Ferocactus Button Brush Rubiaceae Cephalanthus Occidentalis Bottle Brush Myrtaceae Callistemon Esperanza or ‘Yellow Bells’ Bignoniaceae Tecoma Stans Sweetspire Saxifragaceae Itea Virginica Gardenia Rubiaceae Gardenia Hollies Aquifoliaceae Illex Serviceberry Rosaceae Amelanchier ‘Blue Mist’ Spirea Lamiaceae Caryopteris Summer Sweet or Sweet Clethraceae Clethra Alnifolia Pepperbush Flowering Senna Fabaceae Cassia Southern Wax Mrytle Myricaceae Myrica Cerifera Variegated Ligustrum or Oleaceae Ligustrum Privet Sweet Olive Oleaceae Osmanthus Fragrans Scotch Broom Fabaceae Cytisus Scoparius Rose of Sharon Malvaceae Hibiscus (Single Flower) Viburnum Caprifoliaceae Viburnum Spiraea Rosaceae -

Plant Invaders of Mid-Atlantic Natural Areas Revised & Updated – with More Species and Expanded Control Guidance
Plant Invaders of Mid-Atlantic Natural Areas Revised & Updated – with More Species and Expanded Control Guidance National Park Service U.S. Fish and Wildlife Service 1 I N C H E S 2 Plant Invaders of Mid-Atlantic Natural Areas, 4th ed. Authors Jil Swearingen National Park Service National Capital Region Center for Urban Ecology 4598 MacArthur Blvd., N.W. Washington, DC 20007 Britt Slattery, Kathryn Reshetiloff and Susan Zwicker U.S. Fish and Wildlife Service Chesapeake Bay Field Office 177 Admiral Cochrane Dr. Annapolis, MD 21401 Citation Swearingen, J., B. Slattery, K. Reshetiloff, and S. Zwicker. 2010. Plant Invaders of Mid-Atlantic Natural Areas, 4th ed. National Park Service and U.S. Fish and Wildlife Service. Washington, DC. 168pp. 1st edition, 2002 2nd edition, 2004 3rd edition, 2006 4th edition, 2010 1 Acknowledgements Graphic Design and Layout Olivia Kwong, Plant Conservation Alliance & Center for Plant Conservation, Washington, DC Laurie Hewitt, U.S. Fish & Wildlife Service, Chesapeake Bay Field Office, Annapolis, MD Acknowledgements Funding provided by the National Fish and Wildlife Foundation with matching contributions by: Chesapeake Bay Foundation Chesapeake Bay Trust City of Bowie, Maryland Maryland Department of Natural Resources Mid-Atlantic Invasive Plant Council National Capital Area Garden Clubs Plant Conservation Alliance The Nature Conservancy, Maryland–DC Chapter Worcester County, Maryland, Department of Comprehensive Planning Additional Fact Sheet Contributors Laurie Anne Albrecht (jetbead) Peter Bergstrom (European