The Early Middle Miocene Mollusc
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Macrogastra Badia (C. Pfeiffer
Vol. 17(2): 53–62 MACROGASTRA BADIA (C. PFEIFFER, 1828) (GASTROPODA: PULMONATA: CLAUSILIIDAE) IN ZIELENIEC (BYSTRZYCKIE MTS, CENTRAL SUDETES) – ECOLOGY, CONSERVATION STATUS AND LIFE HISTORY – PRELIMINARY DATA TOMASZ K. MALTZ,BEATA M. POKRYSZKO Museum of Natural History, Wroc³aw University, Sienkiewicza 21, 50-335 Wroc³aw, Poland (e-mail: [email protected], [email protected]) ABSTRACT: Information on thedistribution on theAlpine M. badia in Poland dates from the 1960s and was not verified subsequently. A new locality was discovered in 2003 (Bystrzyckie Mts, Zieleniec near Duszniki-Zdrój); it forms a part of a group of isolated, Polish and Czech localities on the border of the species’ distribution range. In the discussed part of the range the species is threatened by habitat destruction and climatic changes. It is legally protected in Poland but preserving its populations requires habitat protection. The preferred habi- tat is herb-rich beech forest, and cool and humid climate is crucial for the species’ survival. The composition of the accompanying malacofauna varies among the sites which is probably associated with their origin. M. badia is oviparous; in May and June it produces batches of 1–3 eggs. The eggs are partly calcified, 1.39–1.61 in major and 1.32–1.45 mm in minor diameter. The incubation period is 16–19 days; the hatching is asynchron- ous; the juveniles reach adult size in 7–8 months. Some data on shell variation are provided; the number of ap- ertural folds varies more widely than formerly believed. KEY WORDS: Clausiliidae, endangered species, Macrogastra badia, lifehistory, ecology INTRODUCTION Macrogastra badia (C. -

Miocene Continental Gastropods from the Southern Margin of the Swabian Alb (Baden-Württemberg, SW Germany)
N. Jb. Geol. Paläont. Abh. 287/1 (2018), 17–44 Article E Stuttgart, January 2018 Miocene continental gastropods from the southern margin of the Swabian Alb (Baden-Württemberg, SW Germany) Olaf Höltke, Rodrigo B. Salvador, and Michael W. Rasser With 6 figures Abstract: The Middle Miocene silvana-beds outcropping at the southern margin of the Swabian Alb bear a wide array of terrestrial and freshwater gastropods. Especially in the surroundings of the Emerberg and Tautschbuch hills, there are records of several collection sites. In the present work, we investigated material from the historical localities from these two regions, housed in museum collec- tions, in order to provide an updated check list, synthesizing the knowledge of the region in the hopes to spur renewed interest. In total, we report 14 freshwater and 50 land snail species from the area, but not all species reported in the literature could be confirmed. Key words: Baden-Württemberg, European Land Mammal Zone MN 5, Langhian, silvana- beds, Upper Freshwater Molasse. 1. Introduction (1923-1930). Therefore, in the absence of more driven excavations for the moment, here we investigated ma- The southern margin of the Swabian Alb, especially terial from several historical localities from these two the Emerberg and Tautschbuch hills, bears a consider- regions, housed in museum collections. Our goal is to able number of continental Miocene deposits (Fig. 1). provide an updated check list of species and localities, These are built up by the silvana-beds, a lithostrati- thus synthesizing the knowledge of the region in the graphic unit of the Miocene Upper Freshwater Molasse hopes to spur renewed interest and collection efforts. -

Pulmonata, Helicidae) and the Systematic Position of Cylindrus Obtusus Based on Nuclear and Mitochondrial DNA Marker Sequences
© 2013 The Authors Accepted on 16 September 2013 Journal of Zoological Systematics and Evolutionary Research Published by Blackwell Verlag GmbH J Zoolog Syst Evol Res doi: 10.1111/jzs.12044 Short Communication 1Centre for Ecological and Evolutionary Synthesis (CEES), University of Oslo, Oslo, Norway; 2Central Research Laboratories, Natural History Museum, Vienna, Austria; 33rd Zoological Department, Natural History Museum, Vienna, Austria; 4Department of Integrative Zoology, University of Vienna, Vienna, Austria; 5Department of Zoology, Hungarian Natural History Museum, Budapest, Hungary New data on the phylogeny of Ariantinae (Pulmonata, Helicidae) and the systematic position of Cylindrus obtusus based on nuclear and mitochondrial DNA marker sequences 1 2,4 2,3 3 2 5 LUIS CADAHIA ,JOSEF HARL ,MICHAEL DUDA ,HELMUT SATTMANN ,LUISE KRUCKENHAUSER ,ZOLTAN FEHER , 2,3,4 2,4 LAURA ZOPP and ELISABETH HARING Abstract The phylogenetic relationships among genera of the subfamily Ariantinae (Pulmonata, Helicidae), especially the sister-group relationship of Cylindrus obtusus, were investigated with three mitochondrial (12S rRNA, 16S rRNA, Cytochrome c oxidase subunit I) and two nuclear marker genes (Histone H4 and H3). Within Ariantinae, C. obtusus stands out because of its aberrant cylindrical shell shape. Here, we present phylogenetic trees based on these five marker sequences and discuss the position of C. obtusus and phylogeographical scenarios in comparison with previously published results. Our results provide strong support for the sister-group relationship between Cylindrus and Arianta confirming previous studies and imply that the split between the two genera is quite old. The tree reveals a phylogeographical pattern of Ariantinae with a well-supported clade comprising the Balkan taxa which is the sister group to a clade with individuals from Alpine localities. -

Liste Rouge Mollusques (Gastéropodes Et Bivalves)
2012 > L’environnement pratique > Listes rouges / Gestion des espèces > Liste rouge Mollusques (gastéropodes et bivalves) Espèces menacées en Suisse, état 2010 > L’environnement pratique > Listes rouges / Gestion des espèces > Liste rouge Mollusques (gastéropodes et bivalves) Espèces menacées en Suisse, état 2010 Publié par l’Office fédéral de l’environnement OFEV et par le Centre suisse de cartographie de la faune CSCF Berne, 2012 Valeur juridique de cette publication Impressum Liste rouge de l’OFEV au sens de l’art. 14, al. 3, de l’ordonnance Editeurs du 16 janvier 1991 sur la protection de la nature et du paysage Office fédéral de l’environnement (OFEV) (OPN; RS 451.1), www.admin.ch/ch/f/rs/45.html L’OFEV est un office du Département fédéral de l’environnement, des transports, de l’énergie et de la communication (DETEC). La présente publication est une aide à l’exécution de l’OFEV en tant Centre Suisse de Cartographie de la Faune (CSCF), Neuchâtel. qu’autorité de surveillance. Destinée en premier lieu aux autorités d’exécution, elle concrétise des notions juridiques indéterminées Auteurs provenant de lois et d’ordonnances et favorise ainsi une application Mollusques terrestres: Jörg Rüetschi, Peter Müller et François Claude uniforme de la législation. Elle aide les autorités d’exécution Mollusques aquatiques: Pascal Stucki et Heinrich Vicentini notamment à évaluer si un biotope doit être considéré comme digne avec la collaboration de Simon Capt et Yves Gonseth (CSCF) de protection (art. 14, al. 3, let. d, OPN). Accompagnement à l’OFEV Francis Cordillot, division Espèces, écosystèmes, paysages Référence bibliographique Rüetschi J., Stucki P., Müller P., Vicentini H., Claude F. -

Malaco Le Journal Électronique De La Malacologie Continentale Française
MalaCo Le journal électronique de la malacologie continentale française www.journal-malaco.fr MalaCo (ISSN 1778-3941) est un journal électronique gratuit, annuel ou bisannuel pour la promotion et la connaissance des mollusques continentaux de la faune de France. Equipe éditoriale Jean-Michel BICHAIN / Paris / [email protected] Xavier CUCHERAT / Audinghen / [email protected] Benoît FONTAINE / Paris / [email protected] Olivier GARGOMINY / Paris / [email protected] Vincent PRIE / Montpellier / [email protected] Les manuscrits sont à envoyer à : Journal MalaCo Muséum national d’Histoire naturelle Equipe de Malacologie Case Postale 051 55, rue Buffon 75005 Paris Ou par Email à [email protected] MalaCo est téléchargeable gratuitement sur le site : http://www.journal-malaco.fr MalaCo (ISSN 1778-3941) est une publication de l’association Caracol Association Caracol Route de Lodève 34700 Saint-Etienne-de-Gourgas JO Association n° 0034 DE 2003 Déclaration en date du 17 juillet 2003 sous le n° 2569 Journal électronique de la malacologie continentale française MalaCo Septembre 2006 ▪ numéro 3 Au total, 119 espèces et sous-espèces de mollusques, dont quatre strictement endémiques, sont recensées dans les différents habitats du Parc naturel du Mercantour (photos Olivier Gargominy, se reporter aux figures 5, 10 et 17 de l’article d’O. Gargominy & Th. Ripken). Sommaire Page 100 Éditorial Page 101 Actualités Page 102 Librairie Page 103 Brèves & News ▪ Endémisme et extinctions : systématique des Endodontidae (Mollusca, Pulmonata) de Rurutu (Iles Australes, Polynésie française) Gabrielle ZIMMERMANN ▪ The first annual meeting of Task-Force-Limax, Bünder Naturmuseum, Chur, Switzerland, 8-10 September, 2006: presentation, outcomes and abstracts Isabel HYMAN ▪ Collecting and transporting living slugs (Pulmonata: Limacidae) Isabel HYMAN ▪ A List of type specimens of land and freshwater molluscs from France present in the national molluscs collection of the Hebrew University of Jerusalem Henk K. -

Molluscs of the Dürrenstein Wilderness Area
Molluscs of the Dürrenstein Wilderness Area S a b i n e F ISCHER & M i c h a e l D UDA Abstract: Research in the Dürrenstein Wilderness Area (DWA) in the southwest of Lower Austria is mainly concerned with the inventory of flora, fauna and habitats, interdisciplinary monitoring and studies on ecological disturbances and process dynamics. During a four-year qualitative study of non-marine molluscs, 96 sites within the DWA and nearby nature reserves were sampled in cooperation with the “Alpine Land Snails Working Group” located at the Natural History Museum of Vienna. Altogether, 84 taxa were recorded (72 land snails, 12 water snails and mussels) including four endemics and seven species listed in the Austrian Red List of Molluscs. A reference collection (empty shells) of molluscs, which is stored at the DWA administration, was created. This project was the first systematic survey of mollusc fauna in the DWA. Further sampling might provide additional information in the future, particularly for Hydrobiidae in springs and caves, where detailed analyses (e.g. anatomical and genetic) are needed. Key words: Wilderness Dürrenstein, Primeval forest, Benign neglect, Non-intervention management, Mollusca, Snails, Alpine endemics. Introduction manifold species living in the wilderness area – many of them “refugees”, whose natural habitats have almost In concordance with the IUCN guidelines, research is disappeared in today’s over-cultivated landscape. mandatory for category I wilderness areas. However, it may not disturb the natural habitats and communities of the nature reserve. Research in the Dürrenstein The Dürrenstein Wilderness Area Wilderness Area (DWA) focuses on providing invento- (DWA) ries of flora and fauna, on interdisciplinary monitoring The Dürrenstein Wilderness Area (DWA) was as well as on ecological disturbances and process dynamics. -

Molluscs, by Michael J
Cambourne New Settlement Iron Age and Romano-British settlement on the clay uplands of west Cambridgeshire Volume 2: Specialist Appendices Web Report 15 Molluscs, by Michael J. Allen Cambourne New Settlement Iron Age and Romano-British Settlement on the Clay Uplands of West Cambridgeshire By James Wright, Matt Leivers, Rachael Seager Smith and Chris J. Stevens with contributions from Michael J. Allen, Phil Andrews, Catherine Barnett, Kayt Brown, Rowena Gale, Sheila Hamilton-Dyer, Kevin Hayward, Grace Perpetua Jones, Jacqueline I. McKinley, Robert Scaife, Nicholas A. Wells and Sarah F. Wyles Illustrations by S.E. James Volume 2: Specialist Appendices Part 1. Artefacts Part 2. Ecofacts Wessex Archaeology Report No. 23 Wessex Archaeology 2009 Published 2009 by Wessex Archaeology Ltd Portway House, Old Sarum Park, Salisbury, SP4 6EB http://www.wessexarch.co.uk Copyright © 2009 Wessex Archaeology Ltd All rights reserved ISBN 978-1-874350-49-1 Project website http://www.wessexarch.co.uk/projects/cambridgeshire/cambourne WA reports web pages http://www.wessexarch.co.uk/projects/cambridgeshire/cambourne/reports ii Contents Web pdf 1 Contents and Concordance of sites and summary details of archive ................................ iii Part 1. Artefacts 2 Prehistoric pottery, by Matt Leivers.....................................................................................1 2 Late Iron Age pottery, by Grace Perpetua Jones................................................................11 2 Romano-British pottery, by Rachael Seager Smith ...........................................................14 -

Fauna of New Zealand Ko Te Aitanga Pepeke O Aotearoa
aua o ew eaa Ko te Aiaga eeke o Aoeaoa IEEAE SYSEMAICS AISOY GOU EESEAIES O ACAE ESEAC ema acae eseac ico Agicuue & Sciece Cee P O o 9 ico ew eaa K Cosy a M-C aiièe acae eseac Mou Ae eseac Cee iae ag 917 Aucka ew eaa EESEAIE O UIESIIES M Emeso eame o Eomoogy & Aima Ecoogy PO o ico Uiesiy ew eaa EESEAIE O MUSEUMS M ama aua Eiome eame Museum o ew eaa e aa ogaewa O o 7 Weigo ew eaa EESEAIE O OESEAS ISIUIOS awece CSIO iisio o Eomoogy GO o 17 Caea Ciy AC 1 Ausaia SEIES EIO AUA O EW EAA M C ua (ecease ue 199 acae eseac Mou Ae eseac Cee iae ag 917 Aucka ew eaa Fauna of New Zealand Ko te Aitanga Pepeke o Aotearoa Number / Nama 38 Naturalised terrestrial Stylommatophora (Mousca Gasooa Gay M ake acae eseac iae ag 317 amio ew eaa 4 Maaaki Whenua Ρ Ε S S ico Caeuy ew eaa 1999 Coyig © acae eseac ew eaa 1999 o a o is wok coee y coyig may e eouce o coie i ay om o y ay meas (gaic eecoic o mecaica icuig oocoyig ecoig aig iomaio eiea sysems o oewise wiou e wie emissio o e uise Caaoguig i uicaio AKE G Μ (Gay Micae 195— auase eesia Syommaooa (Mousca Gasooa / G Μ ake — ico Caeuy Maaaki Weua ess 1999 (aua o ew eaa ISS 111-533 ; o 3 IS -7-93-5 I ie 11 Seies UC 593(931 eae o uIicaio y e seies eio (a comee y eo Cosy usig comue-ase e ocessig ayou scaig a iig a acae eseac M Ae eseac Cee iae ag 917 Aucka ew eaa Māoi summay e y aco uaau Cosuas Weigo uise y Maaaki Weua ess acae eseac O o ico Caeuy Wesie //wwwmwessco/ ie y G i Weigo o coe eoceas eicuaum (ue a eigo oaa (owe (IIusao G M ake oucio o e coou Iaes was ue y e ew eaIa oey oa ue oeies eseac -

Paleoclimatic Reconstruction Using Mutual Climatic Range on Terrestrial Mollusks
Paleoclimatic reconstruction using Mutual Climatic Range on terrestrial mollusks. Olivier Moine, Denis-Didier Rousseau, Dominique Jolly, Marc Vianey-Liaud To cite this version: Olivier Moine, Denis-Didier Rousseau, Dominique Jolly, Marc Vianey-Liaud. Paleoclimatic recon- struction using Mutual Climatic Range on terrestrial mollusks.. Quaternary Research, Elsevier, 2002, 57, pp.162-172. 10.1006/qres.2001.2286. hal-01208190 HAL Id: hal-01208190 https://hal.archives-ouvertes.fr/hal-01208190 Submitted on 2 Oct 2015 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Olivier Moine Paléoenvironnements & Palynologie, Institut des Sciences de l’Evolution (UMR CNRS 5554), Université Montpellier II, case 61, place E. Bataillon, 34095 Montpellier Cedex 5, France Denis-Didier Rousseau Paléoenvironnements & Palynologie, Institut des Sciences de l’Evolution (UMR CNRS 5554), Université Montpellier II, case 61, place E. Bataillon, 34095 Montpellier Cedex 5, France and Lamont-Doherty Earth Observatory of Columbia University, Palisades, New York 10964 Dominique Jolly Paléoenvironnements & Palynologie, Institut des Sciences de l’Evolution (UMR CNRS 5554), Université Montpellier II, case 61, place E. Bataillon, 34095 Montpellier Cedex 5, France Marc Vianey-Liaud Génétique et Environnement, Institut des Sciences de l’Evolution (UMR CNRS 5554), Université Montpellier II, case 64, place E. -
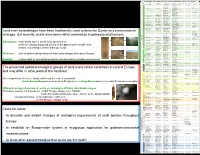
Ecological Groups of Snails – Use and Perspectives
The subdivision of all central European Holocene and Late Glacial land snail species to ecological groups ecological Glacial Early Holocene Middle Holocene Late Holocene (sensu Walker at al 2012) modern immigrants comment group Acanthinula aculeata Acanthinula aculeata Acanthinula aculeata Acanthinula aculeata Acicula parcelineata Acicula parcelineata Aegopinella epipedostoma one sites Aegopinella nitens Aegopinella nitens Aegopinella nitidula Aegopinella nitidula few sites Aegopinella pura Aegopinella pura Aegopinella pura Aegopinella pura Aegopis verticillus Ecological groups of snails Argna bielzi Argna bielzi Bulgarica cana Bulgarica cana Carpathica calophana Carpathica calophana one site; undated Causa holosericea Causa holosericea Clausilia bidentata no fossil data Clausilia cruciata Clausilia cruciata Clausilia cruciata – use and perspectives Cochlodina laminata Cochlodina laminata Cochlodina laminata Cochlodina laminata Cochlodina orthostoma Cochlodina orthostoma Cochlodina orthostoma Cochlodina orthostoma Daudebardia brevipes Daudebardia brevipes Daudebardia rufa Daudebardia rufa Daudebardia rufa Daudebardia rufa Discus perspectivus Discus perspectivus Discus perspectivus 1 2 1 1 ) Lucie Juřičková , Michal Horsák , Jitka Horáčková and Vojen Ložek Discus ruderatus Discus ruderatus Discus ruderatus Discus ruderatus Ena montana Ena montana Ena montana Ena montana forest Eucobresia nivalis Eucobresia nivalis Eucobresia nivalis Faustina faustina Faustina faustina Faustina faustina Faustina faustina Faustina rossmaessleri Faustina -

T.C. Süleyman Demirel Üniversitesi Fen Bilimleri
T.C. SÜLEYMAN DEM İREL ÜN İVERS İTES İ FEN B İLİMLER İ ENST İTÜSÜ KUZEYBATI ANADOLU’NUN KARASAL GASTROPODLARI ÜM İT KEBAPÇI Danı şman: Prof. Dr. M. Zeki YILDIRIM DOKTORA TEZ İ BİYOLOJ İ ANAB İLİMDALI ISPARTA – 2007 Fen Bilimleri Enstitüsü Müdürlü ğüne Bu çalı şma jürimiz tarafından …………. ANAB İLİM DALI'nda oybirli ği/oyçoklu ğu ile DOKTORA TEZ İ olarak kabul edilmi ştir. Ba şkan : (Ünvanı, Adı ve Soyadı) (İmza) (Kurumu)................................................... Üye : (Ünvanı, Adı ve Soyadı) (İmza) (Kurumu)................................................... Üye : (Ünvanı, Adı ve Soyadı) (İmza) (Kurumu)................................................... Üye: (Ünvanı, Adı ve Soyadı) (İmza) (Kurumu)................................................... Üye : (Ünvanı, Adı ve Soyadı) (İmza) (Kurumu)................................................... ONAY Bu tez .../.../20.. tarihinde yapılan tez savunma sınavı sonucunda, yukarıdaki jüri üyeleri tarafından kabul edilmi ştir. ...../...../20... Prof. Dr. Fatma GÖKTEPE Enstitü Müdürü İÇİNDEK İLER Sayfa İÇİNDEK İLER......................................................................................................... i ÖZET........................................................................................................................ ix ABSTRACT.............................................................................................................. x TE ŞEKKÜR ............................................................................................................. xi ŞEK -

Fauna of New Zealand Website Copy 2010, Fnz.Landcareresearch.Co.Nz
aua o ew eaa Ko te Aiaga eeke o Aoeaoa IEEAE SYSEMAICS AISOY GOU EESEAIES O ACAE ESEAC ema acae eseac ico Agicuue & Sciece Cee P O o 9 ico ew eaa K Cosy a M-C aiièe acae eseac Mou Ae eseac Cee iae ag 917 Aucka ew eaa EESEAIE O UIESIIES M Emeso eame o Eomoogy & Aima Ecoogy PO o ico Uiesiy ew eaa EESEAIE O MUSEUMS M ama aua Eiome eame Museum o ew eaa e aa ogaewa O o 7 Weigo ew eaa EESEAIE O OESEAS ISIUIOS awece CSIO iisio o Eomoogy GO o 17 Caea Ciy AC 1 Ausaia SEIES EIO AUA O EW EAA M C ua (ecease ue 199 acae eseac Mou Ae eseac Cee iae ag 917 Aucka ew eaa Fauna of New Zealand Ko te Aitanga Pepeke o Aotearoa Number / Nama 38 Naturalised terrestrial Stylommatophora (Mousca Gasooa Gay M ake acae eseac iae ag 317 amio ew eaa 4 Maaaki Whenua Ρ Ε S S ico Caeuy ew eaa 1999 Coyig © acae eseac ew eaa 1999 o a o is wok coee y coyig may e eouce o coie i ay om o y ay meas (gaic eecoic o mecaica icuig oocoyig ecoig aig iomaio eiea sysems o oewise wiou e wie emissio o e uise Caaoguig i uicaio AKE G Μ (Gay Micae 195— auase eesia Syommaooa (Mousca Gasooa / G Μ ake — ico Caeuy Maaaki Weua ess 1999 (aua o ew eaa ISS 111-533 ; o 3 IS -7-93-5 I ie 11 Seies UC 593(931 eae o uIicaio y e seies eio (a comee y eo Cosy usig comue-ase e ocessig ayou scaig a iig a acae eseac M Ae eseac Cee iae ag 917 Aucka ew eaa Māoi summay e y aco uaau Cosuas Weigo uise y Maaaki Weua ess acae eseac O o ico Caeuy Wesie //wwwmwessco/ ie y G i Weigo o coe eoceas eicuaum (ue a eigo oaa (owe (IIusao G M ake oucio o e coou Iaes was ue y e ew eaIa oey oa ue oeies eseac