Integrating Climate Change Into Northeast and Midwest State Wildlife Action Plans
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
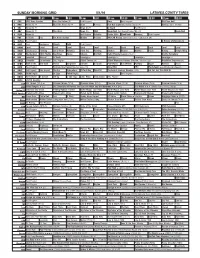
Sunday Morning Grid 5/1/16 Latimes.Com/Tv Times
SUNDAY MORNING GRID 5/1/16 LATIMES.COM/TV TIMES 7 am 7:30 8 am 8:30 9 am 9:30 10 am 10:30 11 am 11:30 12 pm 12:30 2 CBS CBS News Sunday Face the Nation (N) Paid Program Boss Paid Program PGA Tour Golf 4 NBC News (N) Å Meet the Press (N) Å News Rescue Red Bull Signature Series (Taped) Å Hockey: Blues at Stars 5 CW News (N) Å News (N) Å In Touch Paid Program 7 ABC News (N) Å This Week News (N) NBA Basketball First Round: Teams TBA. (N) Basketball 9 KCAL News (N) Joel Osteen Schuller Pastor Mike Woodlands Amazing Paid Program 11 FOX In Touch Paid Fox News Sunday Midday Prerace NASCAR Racing Sprint Cup Series: GEICO 500. (N) 13 MyNet Paid Program A History of Violence (R) 18 KSCI Paid Hormones Church Faith Paid Program 22 KWHY Local Local Local Local Local Local Local Local Local Local Local Local 24 KVCR Landscapes Painting Joy of Paint Wyland’s Paint This Painting Kitchen Mexico Martha Pépin Baking Simply Ming 28 KCET Wunderkind 1001 Nights Bug Bites Space Edisons Biz Kid$ Celtic Thunder Legacy (TVG) Å Soulful Symphony 30 ION Jeremiah Youssef In Touch Leverage Å Leverage Å Leverage Å Leverage Å 34 KMEX Conexión En contacto Paid Program Fútbol Central (N) Fútbol Mexicano Primera División: Toluca vs Azul República Deportiva (N) 40 KTBN Walk in the Win Walk Prince Carpenter Schuller In Touch PowerPoint It Is Written Pathway Super Kelinda Jesse 46 KFTR Paid Program Formula One Racing Russian Grand Prix. -

INGO GILDENHARD Cicero, Philippic 2, 44–50, 78–92, 100–119 Latin Text, Study Aids with Vocabulary, and Commentary CICERO, PHILIPPIC 2, 44–50, 78–92, 100–119
INGO GILDENHARD Cicero, Philippic 2, 44–50, 78–92, 100–119 Latin text, study aids with vocabulary, and commentary CICERO, PHILIPPIC 2, 44–50, 78–92, 100–119 Cicero, Philippic 2, 44–50, 78–92, 100–119 Latin text, study aids with vocabulary, and commentary Ingo Gildenhard https://www.openbookpublishers.com © 2018 Ingo Gildenhard The text of this work is licensed under a Creative Commons Attribution 4.0 International license (CC BY 4.0). This license allows you to share, copy, distribute and transmit the text; to adapt the text and to make commercial use of the text providing attribution is made to the author(s), but not in any way that suggests that they endorse you or your use of the work. Attribution should include the following information: Ingo Gildenhard, Cicero, Philippic 2, 44–50, 78–92, 100–119. Latin Text, Study Aids with Vocabulary, and Commentary. Cambridge, UK: Open Book Publishers, 2018. https://doi. org/10.11647/OBP.0156 Every effort has been made to identify and contact copyright holders and any omission or error will be corrected if notification is made to the publisher. In order to access detailed and updated information on the license, please visit https:// www.openbookpublishers.com/product/845#copyright Further details about CC BY licenses are available at http://creativecommons.org/licenses/ by/4.0/ All external links were active at the time of publication unless otherwise stated and have been archived via the Internet Archive Wayback Machine at https://archive.org/web Digital material and resources associated with this volume are available at https://www. -

Historical Painting Techniques, Materials, and Studio Practice
Historical Painting Techniques, Materials, and Studio Practice PUBLICATIONS COORDINATION: Dinah Berland EDITING & PRODUCTION COORDINATION: Corinne Lightweaver EDITORIAL CONSULTATION: Jo Hill COVER DESIGN: Jackie Gallagher-Lange PRODUCTION & PRINTING: Allen Press, Inc., Lawrence, Kansas SYMPOSIUM ORGANIZERS: Erma Hermens, Art History Institute of the University of Leiden Marja Peek, Central Research Laboratory for Objects of Art and Science, Amsterdam © 1995 by The J. Paul Getty Trust All rights reserved Printed in the United States of America ISBN 0-89236-322-3 The Getty Conservation Institute is committed to the preservation of cultural heritage worldwide. The Institute seeks to advance scientiRc knowledge and professional practice and to raise public awareness of conservation. Through research, training, documentation, exchange of information, and ReId projects, the Institute addresses issues related to the conservation of museum objects and archival collections, archaeological monuments and sites, and historic bUildings and cities. The Institute is an operating program of the J. Paul Getty Trust. COVER ILLUSTRATION Gherardo Cibo, "Colchico," folio 17r of Herbarium, ca. 1570. Courtesy of the British Library. FRONTISPIECE Detail from Jan Baptiste Collaert, Color Olivi, 1566-1628. After Johannes Stradanus. Courtesy of the Rijksmuseum-Stichting, Amsterdam. Library of Congress Cataloguing-in-Publication Data Historical painting techniques, materials, and studio practice : preprints of a symposium [held at] University of Leiden, the Netherlands, 26-29 June 1995/ edited by Arie Wallert, Erma Hermens, and Marja Peek. p. cm. Includes bibliographical references. ISBN 0-89236-322-3 (pbk.) 1. Painting-Techniques-Congresses. 2. Artists' materials- -Congresses. 3. Polychromy-Congresses. I. Wallert, Arie, 1950- II. Hermens, Erma, 1958- . III. Peek, Marja, 1961- ND1500.H57 1995 751' .09-dc20 95-9805 CIP Second printing 1996 iv Contents vii Foreword viii Preface 1 Leslie A. -

Status Report from Dutch AAT, Weda, 2017
Update Dutch AAT ITWG 2017 Content . Translation work . Management software Xtree . Concept proposals . Visual fashion and costume thesaurus . Events 6420 new concepts (2012-2016) Pref terms: 6420 Alternative terms: 1917 Use for terms: 535 Translation labels Translation N/A: 1598 literal translation: 749 Loanword: 143 Translation work Masaai (language) http://vocab.getty.edu/page/aat/300388758 Vitales (genus): http://vocab.getty.edu/page/aat/300389726 Tefon™: http://vocab.getty.edu/page/aat/300380055 Translation NA Abandoned dwellings/ verlaten woningen: http://vocab.getty.edu/page/aat/300386972 Singel sitter models/ portretten met één model http://vocab.getty.edu/page/aat/300404138 suminagashi paper/ suminagashi-papier http://vocab.getty.edu/page/aat/300387534 Literal translation Katana: http://vocab.getty.edu/page/aat/300386050 Buon Fresco http://vocab.getty.edu/page/aat/300403944 abhaya mudra: http://vocab.getty.edu/page/aat/300386135 Loanwords Xtree thesaurus management Software What is xTree? • A central, web-based tool to administer vocabularies (e.g. thesauri, classifications, lists, taxonomies, folksonomies) • is semantically interoperable within ISO/DIS 25964-1 (ThesaurusConcept, ThesaurusArray, ConceptGroup), ANSI/NISO Z39.19-2005, ISO 2788 and DIN 1463-1 • is compatible with the data model of BS DD 8723-5 • has a multi-lingual data model • is usable with common web-browser software (e.g. Firefox 3, Internet Explorer 8) • is easy to handle (e.g. drag and drop) • uses the museumvok Webservice • uses SKOS RDF • is built for collaborative work http://xtree-public.digicult- • is developed with Open Source software verbund.de/vocnet/? • XSD- and RDF-based Webservice Xtree, managentsoftware New flag: ‘translation N/A’ Dutch AAT has a set of ‘Untranslatables’ How to flag these concepts? Checked but not translated… ‘half-tester beds’ ID 300038710 New flag: ‘literalN.B. -

JPEG Image Compression2.Pdf
JPEG image compression FAQ, part 2/2 2/18/05 5:03 PM Part1 - Part2 - MultiPage JPEG image compression FAQ, part 2/2 There are reader questions on this topic! Help others by sharing your knowledge Newsgroups: comp.graphics.misc, comp.infosystems.www.authoring.images From: [email protected] (Tom Lane) Subject: JPEG image compression FAQ, part 2/2 Message-ID: <[email protected]> Summary: System-specific hints and program recommendations for JPEG images Keywords: JPEG, image compression, FAQ, JPG, JFIF Reply-To: [email protected] Date: Mon, 29 Mar 1999 02:24:34 GMT Sender: [email protected] Archive-name: jpeg-faq/part2 Posting-Frequency: every 14 days Last-modified: 28 March 1999 This article answers Frequently Asked Questions about JPEG image compression. This is part 2, covering system-specific hints and program recommendations for a variety of computer systems. Part 1 covers general questions and answers about JPEG. As always, suggestions for improvement of this FAQ are welcome. New since version of 14 March 1999: * Added entries for PIE (Windows digicam utility) and Cameraid (Macintosh digicam utility). * New version of VuePrint (7.3). This article includes the following sections: General info: [1] What is covered in this FAQ? [2] How do I retrieve these programs? Programs and hints for specific systems: [3] X Windows [4] Unix (without X) [5] MS-DOS [6] Microsoft Windows [7] OS/2 [8] Macintosh [9] Amiga [10] Atari ST [11] Acorn Archimedes [12] NeXT [13] Tcl/Tk [14] Other systems Source code for JPEG: [15] Freely available source code for JPEG Miscellaneous: [16] Which programs support progressive JPEG? [17] Where are FAQ lists archived? This article and its companion are posted every 2 weeks. -

Installation Manual ESPAÑOL Instrucciones De Instalación FRANÇAIS Instructions D’Installation
SUNNY BOY 3.0-US / 3.8-US / 5.0-US / 6.0-US / 7.0-US / 7.7-US SB3.0-1SP-US-41 / SB3.8-1SP-US-41 / SB5.0-1SP-US-41 / SB6.0-1SP-US-41 / SB7.0-1SP-US-41 / SB7.7-1SP-US-41 SUNNY BO Y eManual ENGLISH Installation Manual ESPAÑOL Instrucciones de instalación FRANÇAIS Instructions d’installation SBxx-1SP-US-41-IA-xx-10 | 117821-00.01 | Version 1.0 Legal Provisions SMA Solar Technology AG ENGLISH Legal Provisions The information contained in these documents is the property of SMA Solar Technology AG. No part of this document may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, be it electronic, mechanical, photographic, magnetic or otherwise, without the prior written permission of SMA Solar Technology AG. Internal reproduction used solely for the purpose of product evaluation or other proper use is allowed and does not require prior approval. SMA Solar Technology AG makes no representations or warranties, express or implied, with respect to this documentation or any of the equipment and/or software it may describe, including (with no limitation) any implied warranties of utility, merchantability, or fitness for any particular purpose. All such representations or warranties are expressly disclaimed. Neither SMA Solar Technology AG nor its distributors or dealers shall be liable for any indirect, incidental, or consequential damages under any circumstances. The exclusion of implied warranties may not apply in all cases under some statutes, and thus the above exclusion may not apply. -

Honoring America's Fallen Heroes: An
HONORING AMERICA’S FALLEN HEROES: AN UPDATE ON OUR NATIONAL CEMETERIES HEARING BEFORE THE SUBCOMMITTEE ON DISABILITY ASSISTANCE AND MEMORIAL AFFAIRS OF THE COMMITTEE ON VETERANS’ AFFAIRS U.S. HOUSE OF REPRESENTATIVES ONE HUNDRED TWELFTH CONGRESS SECOND SESSION THURSDAY, MARCH 8, 2012 Serial No. 112–50 Printed for the use of the Committee on Veterans’ Affairs ( U.S. GOVERNMENT PRINTING OFFICE 73–294 WASHINGTON : 2013 For sale by the Superintendent of Documents, U.S. Government Printing Office Internet: bookstore.gpo.gov Phone: toll free (866) 512–1800; DC area (202) 512–1800 Fax: (202) 512–2104 Mail: Stop IDCC, Washington, DC 20402–0001 VerDate Aug 31 2005 08:01 Apr 16, 2013 Jkt 000000 PO 00000 Frm 00001 Fmt 5011 Sfmt 5011 Y:\112CONG\DAMA\3-8-12\GPO\73294.TXT LEN VACREP180 with DISTILLER COMMITTEE ON VETERANS’ AFFAIRS JEFF MILLER, Florida, Chairman CLIFF STEARNS, Florida BOB FILNER, California, Ranking DOUG LAMBORN, Colorado CORRINE BROWN, Florida GUS M. BILIRAKIS, Florida SILVESTRE REYES, Texas DAVID P. ROE, Tennessee MICHAEL H. MICHAUD, Maine MARLIN A. STUTZMAN, Indiana LINDA T. SA´ NCHEZ, California BILL FLORES, Texas BRUCE L. BRALEY, Iowa BILL JOHNSON, Ohio JERRY McNERNEY, California JEFF DENHAM, California JOE DONNELLY, Indiana JON RUNYAN, New Jersey TIMOTHY J. WALZ, Minnesota DAN BENISHEK, Michigan JOHN BARROW, Georgia ANN MARIE BUERKLE, New York RUSS CARNAHAN, Missouri TIM HUELSKAMP, Kansas MARK E. AMODEI, Nevada ROBERT L. TURNER, New York HELEN W. TOLAR, Staff Director and Chief Counsel SUBCOMMITTEE ON DISABILITY ASSISTANCE AND MEMORIAL AFFAIRS JON RUNYAN, New Jersey, Chairman DOUG LAMBORN, Colorado JERRY McNERNEY, California, Ranking ANN MARIE BUERKLE, New York JOHN BARROW, Georgia MARLIN A. -

2101-Washington-Ave-Nomination.Pdf
NOMINATION OF HISTORIC B UILDING , S TRUCTURE , S ITE , OR OBJECT P HILADELPHIA R EGISTER OF HISTORIC P LACES P HILADELPHIA HISTORICAL C OMMISSION SUBMIT ALL ATTACHED MATERIALS ON PAPER AND IN ELECTRONIC FORM ON CD (M S WORD FORMAT) 1. ADDRESS OF HISTORIC RESOURCE (must comply with an Office of Property Assessment address) Street address:_____________2101____ _Washington_________ _Avenue______________________________________ Postal code:__19146-2532_____________ Councilmanic District:________2__________________ 2. NAME OF HISTORIC RESOURCE Historic Name:______Howell______ &_ _Brothers_______ Wallpaper________ _Hangings_______ _Manufactory____________________________ Common Name:________Frankford_______ _Chocolate________ Company________ _Building________________________________ 3. TYPE OF HISTORIC R ESOURCE X Building Structure Site Object 4. P ROPERTY INFORMATION Condition: excellent good X fair poor ruins Occupancy: occupied X vacant under construction unknown Current use:__________Telecommunications_________________ and___ _Automobile_________ _Parking___________________________ 5. BOUNDARY DESCRIPTION See Page 1 6. DESCRIPTION See Page 2 7. S IGNIFICANCE See Page 13 Period of Significance (from year to year): from __1865_______ to __2012_______ Date(s) of construction and/or alteration:_ _1865____, _1883______, 1912_______________________, 1964, 1967, 1968, 1975__________ 1865, 83: Unknown 1912: William C Haddock 1964: Uknown 1967, 68: Abraham Levy Architect, engineer, and/or designer:__1975:___ Unknown____________________________________________ -

Heritage Studies: an Odyssey in Learning
Heritage Studies: An Odyssey in Learning The Heritage Guide 2005-2006 1 TABLE OF CONTENTS I. INTRODUCTION ..................................................................... 3 II. HERITAGE TEXTS ................................................................. 6 III. HERITAGE SEMINARS Heritage I .......................................................................... 7 Heritage II ......................................................................... 16 Heritage III: Asia............................................................... 29 Heritage III: Latin America............................................... 39 IV. GOALS AND OBJECTIVES Reading ............................................................................. 46 Writing .............................................................................. 47 Cultural Literacy ............................................................... 49 Oral Communication ........................................................ 50 V. CLASS REQUIREMENTS AND EXPECTATIONS Attendance………………………..................................... 51 Registering for a Class; Add/Drop Procedures................. 51 How to Protect Your Work............................................... 51 Academic Honesty and Plagiarism.................................... 52 Penalties for Plagiarism..................................................... 53 How to Avoid Plagiarism.................................................. 53 VI. READING, WRITING, SPEAKING, AND LISTENING IN HERITAGE Reading ............................................................................ -

National Petroleum Reserve-Alaska Final Integrated Activity Plan/Environmental Impact Statement I Contents
National Petroleum Reserve-Alaska FINAL Integrated Activity Plan/ Environmental Impact Statement Volume 6 Appendices, Glossary, Bibliography Prepared by: U.S. Department of the Interior Bureau of Land Management Anchorage, Alaska In cooperation with: North Slope Borough U.S. Bureau of Ocean Energy Management U.S. Fish and Wildlife Service November 2012 How the IAP/EIS is Organized VOLUME 1 Chapter 1 – Introduction: Summarizes the purpose of and need for this IAP/EIS and decisions to be made. Chapter 2 – Alternatives: Describes and compares proposed management alternatives. Chapter 3 – Affected Environment: Presents existing natural and socioeconomic resources in the NPR-A and trends, including those associated with climate change. VOLUME 2 Chapter 4 – Environmental Consequences (sections 4.1 – 4.4): Provides the assumptions upon which the impact analysis rests and evaluates impacts of Alternatives A and B-1 on resources and uses in the NPR-A relevant to making a decision among the alternatives. VOLUME 3 Chapter 4 – Environmental Consequences continued (sections 4.5 – 4.7): Evaluates impacts of Alternatives B-2 (preferred alternative), C, and D on resources and uses in the NPR-A relevant to making a decision among the alternatives. VOLUME 4 Chapter 4 – Environmental Consequences continued (sections 4.8 – 4.13): Evaluates the cumulative impacts on resources and uses in the NPR-A and other effects relevant to making a decision among the alternatives. VOLUME 5 Chapter 5 – Consultation and Coordination: Describes public and government (including tribal) consultation undertaken for this plan and the development of alternatives and lists the plan’s preparers. Chapter 6 – Comments and Responses: Presents public comments on the Draft IAP/EIS and responses to the comments. -
Newcomers Claim Three Council Seats
THREE DAYS A WEEK POST COMMENTS AT CAPE-CORAL-DAILY-BREEZE.COM Season CAPE CORAL preview Local soccer squads getting ready for openers BREEZE — SPORTS MID-WEEK EDITION WEATHER: Partly Sunny • Tonight: Mostly Clear • Friday: Mostly Cloudy — 2A cape-coral-daily-breeze.com Vol. 50, No. 135 Thursday, November 10, 2011 50 cents Newcomers claim three council seats of the vote; and Donnell won over “I don’t think they’re doing One incumbent wins re-election, one amendment approved Stokes with 51.25 percent of the what they’re supposed to do,” By DREW WINCHESTER bloc, of city council for at least “Scott” Morris to claim the four vote. Cole said. “They’re thinking of [email protected] the next two years. available seats. Joyce Cole, a 35-year Cape themselves and not the people or Two incumbent council mem- Council members Pete Brandt Carioscia beat Brandt in resident, said recent actions by the city as a whole.” bers were shown the door by vot- and Bill Deile were bested by District 2 with 56.90 percent of council members and City For some voters, like Doris ers while one survived on challengers John Carioscia and the vote; Nesta topped Deile in Manager Gary King, in particular Schroeder, the election came Tuesday night, joining one local Lenny Nesta, while Dr. Derrick District 3 with 55.46 percent of his desire for an added bonus to down to a single issue. Schroeder political newcomer to change the Donnell beat David Stokes and the vote; Erbrick took the race his salary, tipped the scale in make-up, and no doubt the voting Rana Erbrick outlasted WIlliam from Morris with 53.45 percent favor of the challengers. -

Artist Citations
Downtown Fremont Public Art Program (Temporary Installation) – Metamorphosis Downloaded by Alina Kwak Thursday, December 15th 2016, 2:27pm Call ID: 990676 Artist ID: 7754 Status: Received Helle scharlingtodd Contact Partner Contact via Email [email protected] Phone Cell Web Site glassandmosaics.com Mailing Address . Custom Answers 2) Statement Regarding ThemeArtist must provide a written statement (up to 150 words) as part of their submittal explaining how their sculpture addresses and illustrates the theme. Art is like music, it stimulates the mind and the senses. With this in mind I build my pieces with emphasis on space, form, color, line, rhythm, and content. Artists are image makers and like the world's antennas, they feel, think and express reality. Momentarily I focus on "defining our spaces", like building a new society, using color, form and line as the building blocks. In addition to my public art projects I have developed a series of "color molecule" sculptures. The sculptures represent an homage to the smallest building block of life, the molecules. The theme is called "metamorphosis" which means change, and I see my work fitiing to the theme, as we all have metamorphed from molecules to a human being, so honoring the smallest entity in us seems allright. The work has semi circular motions, which gives a light feeling. The piece is 14' x 6;' x 6'. It is powder coated, the main color being turquise with colored glass inserted in the molecules. When the light shines through 3) Art Installation and De-Installation PlanPlease reference the attached site plan and provide an installation and de-installation plan and requirements for the attachment of the Artwork and the type of hardware to be used for the attachment.