Flower Characteristics, VOC Emission Profile, and Glandular
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Bees and Wasps of Marsland Nature Reserve
The Bees and Wasps of Marsland Nature Reserve Mason wasp Invertebrate survey and habitat evaluation Patrick Saunders [email protected] http://kernowecology.co.uk 1 Introduction This document consists of habitat evaluation and management recommendations for Bees and Wasps (Aculeate hymenoptera) for the Devon Wildlife Trust Nature Reserve Marsland mouth. The survey and report was commissioned by DWT Reserve warden. Marsland Nature reserve description (Pilkington & Threlkeld 2012) • The reserve comprises 212 hectares, of which 186 hectares occurs in the Marsland Valley and 26 hectares in the Welcombe Valley. The site was designated a SSSI in 1952. In addition the reserve includes an unknown acreage of foreshore north of Welcombe Mouth for 4 kilometres, extending beyond South Hole Farm (SS219201). The boundary of the reserve is approximately 18 miles long and is very complex, mainly through following the seven separate tributary streams. The reserve is freehold owned by Devon Wildlife Trust • The primary interest of the reserve is as an example of a north Devon/Cornwall coombe valley with a variety of slopes, soil types and aspects and coastal area that gives rise to a similar diversity of habitats. The most important of these are the extensive areas of relatively pure oak woodland and oak coppice, the maritime grassland and grass heath and the alder woodland and wet flushes in the valley bottoms. • There is approximately 36h of grassland, 130h of woodland, 43h of coastal habitat and 1h of open water. • The reserve also lies within an Area of Outstanding Natural Beauty with the Marsland Valley being highly representative of an unspoilt coastal coombe habitat. -

Chemodiversity of Exudate Flavonoids in Some Members of the Lamiaceae
Biochemical Systematics and Ecology 31 (2003) 1279–1289 www.elsevier.com/locate/biochemsyseco Chemodiversity of exudate flavonoids in some members of the Lamiaceae Karin M. Valant-Vetschera a,∗, James N. Roitman b, Eckhard Wollenweber c a Institut fu¨r Botanik der Universita¨t Wien, Rennweg 14, A-1030 Vienna, Austria b Plant Mycotoxin Research, USDA–ARS–Western Regional Research Center, 800 Buchanan Street, Albany, CA 94710, USA c Institut fu¨r Botanik der TU Darmstadt, Schnittspahnstraße 3, D-64287 Darmstadt, Germany Received 30 August 2002; accepted 3 January 2003 Abstract Several newly studied species and further accessions of the Lamiaceae have been analyzed for their exudate flavonoid profiles. The principal compounds accumulated were flavones and their 6-methoxy derivatives, whereas flavonols were rarely encountered. The chemodiversity observed was relatively low, with only some 15 derivatives being found. The new data are discussed in relation to published data, and chemosystematic aspects are briefly addressed. Of the studied species, Salvia arizonica yielded only a rare diterpene quinone, demethylfruticulin A. Glandular hair diversification and different qualities of their secretions are briefly discussed. 2003 Elsevier Science Ltd. All rights reserved. Keywords: Teucrium; Salvia; Phlomis; Dorystoechas; Lamiaceae; Exudate flavonoids; Diterpene quinone; Chemodiversity; Chemosystematics 1. Introduction The family of Lamiaceae consists of approximately 200 genera of cosmopolitan distribution, many of them of economic importance due to essential oil production. Most genera of the Lamiaceae are thus rich sources of terpenoids, but in addition a variety of iridoid glycosides and flavonoids is accumulated in considerable amount ∗ Corresponding author. Tel.: +43-1-4277-54102; fax: +43-1-4277-9541. -

Essential Oil Composition of Teucrium L. (Lamiaceae) Collected from Different Locations in Libya
THE THIRD INTERNATIONAL CONFERENCE ON BASIC SCIENCES & THEIR APPLICATIONS Code: Bota 115 Essential oil composition of Teucrium L. (Lamiaceae) collected from different locations in Libya. Abdelbaset M.Asker*1, Salem A.Hassan2 and Baset E.S.Mohammed3 1Botany Department, Faculty of Science, Omar Al Mukhtar University, Al Baida, Libya. 2Bioilogy Departmen( Botany ), Faculty of Science,Al-Margeb University 3School of Education,Biological Science, Omar Al Mukhtar University, Ghubah, Libya. Corresponding author: [email protected] ABSTRACT A The essential oil was extracted from the dried shoots of plant materials, twelve compounds were identified by Gas chromatography–mass spectrometry (GC-MS) analyses revealed that essential oil contains mainly germacrene B, β-caryophyllene, Limonene, α-pinene, Germacrene D, β-Elemene, α-Copaene, α-Cadinol, Terpinen, Isoborneol, Camphene and Linalool. However, T. zanonii was specified by the presence of all assessed oils, while section Chamaedrys (T.barbeyanum) was characterized by the absence of α-pinene, α-cadinol and Isoborneol. Although, α- pinene and α-cadinol were also absent in T.polium subsp. flavovirens, these were detected in the two other forms of T.polium. Contrary, Linalool and Terpinen were detected in the two forms of T.polium and not detect in subspecies. The Terpinen is also undetected in T.brevifolium, T.campanulatum and T.fruticans, which are belonged to section Teucrium. Keywords: Teucrium, Lamiaceae Essential oil, GC/MS, Libya. Introduction Lamiaceae is a cosmopolitan family with more than containing about 236 genera and has been stated to contain 6900–7200 species [1]. Fifty percent of the known species restricted to ten genera; Clerodendrum, Hyptis, Nepeta, Plectranthus, Scutellaria, Salvia, Stachys, Thymus, Teucrium, and Vitex. -

Potentials of Pesticidal Plants in Enhancing Diversity of Pollinators in Cropped Fields
American Journal of Plant Sciences, 2018, 9, 2659-2675 http://www.scirp.org/journal/ajps ISSN Online: 2158-2750 ISSN Print: 2158-2742 Potentials of Pesticidal Plants in Enhancing Diversity of Pollinators in Cropped Fields Juliana Godifrey*, Ernest R. Mbega, Patrick A. Ndakidemi Department of Sustainable Agriculture and Biodiversity Ecosystems Management School of Life Science and Bio-Engineering, The Nelson Mandela African Institution of Science and Technology (NM-AIST), Arusha, Tanzania How to cite this paper: Godifrey, J., Mbe- Abstract ga, E.R. and Ndakidemi, P.A. (2018) Poten- tials of Pesticidal Plants in Enhancing Di- Declines in populations of pollinators in agricultural based landscapes have versity of Pollinators in Cropped Fields. raised a concern, which could be associated with various factors such as in- American Journal of Plant Sciences, 9, tensive farming systems like monocropping and the use of non-selective syn- 2659-2675. thetic pesticides. Such practices are likely to remove beneficial non-crop https://doi.org/10.4236/ajps.2018.913193 plants around or nearby the cropped fields. This may in turn result into losses Received: September 25, 2018 of pollinators due to loss of the natural habitats for insects therefore, inter- Accepted: December 18, 2018 fering the interaction between beneficial insects and flowering crop plants. Published: December 21, 2018 Initiatives to restore friendly habitats for pollinators require multidisciplinary approaches. One of these could be the use of pesticidal flowering plants as Copyright © 2018 by authors and part of field margin plants with the aim of encouraging the population of pol- Scientific Research Publishing Inc. This work is licensed under the Creative linators whilst reducing the number of pests. -

How to Attract and Maintain Pollinators in Your Garden MARISSA V
ANR Publication 8498 | October 2013 http://anrcatalog.ucanr.edu How to Attract and Maintain Pollinators in Your Garden MARISSA V. PONDER, Laboratory NTRODUCTION Assistant, University of California, I Berkeley; GORDON W. FRANKIE, Entomologist, University of early all ecosystems on earth depend on pollination California, Berkeley; RACHEL Nof flowering plants for their existence and survival; ELKINS, UC Cooperative Extension furthermore, from 70 to 75 percent of the world’s Pomology Advisor, Lake and flowering plants and over one-third of the world’s crop Mendocino Counties; KATE FREY, International Landscape Designer; species depend on pollination for reproduction (Klein ROLLIN COVILLE, Photographer, et al. 2007; NAS 2007). Take a stroll through your University of California, Berkeley; neighborhood or a botanical garden, or hike in the MARY SCHINDLER, Laboratory hills, and experience the shapes and smells of flowers Assistant, University of California, surrounding you. When most people look at a flower, Berkeley; SARA LEON GUERRERO, Laboratory Assistant, University of they notice the shape, smell, composition, or structure California, Berkeley; of the flower, but few take a moment to consider why JAIME C. PAWELEK, Laboratory the blossom appears and smells as it does (Frey 2001). Assistant, University of California, Plants have evolved through time to offer unique flowers Berkeley; and CAROLYN SHAFFER, that attract select pollinators, thus ensuring that the Laboratory Assistant, UC Cooperative Extension, pollinator’s visits will provide them with another year of Lake County flowers and fruiting. The end result of the pollination process is that humans and animals of all kinds benefit from a bountiful supply of food and beauty (NAS 2007). -
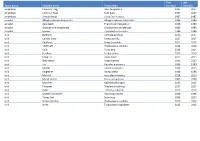
Taxon Group Common Name Taxon Name First Recorded Last
First Last Taxon group Common name Taxon name recorded recorded amphibian Common Frog Rana temporaria 1987 2017 amphibian Common Toad Bufo bufo 1987 2017 amphibian Smooth Newt Lissotriton vulgaris 1987 1987 annelid Alboglossiphonia heteroclita Alboglossiphonia heteroclita 1986 1986 annelid duck leech Theromyzon tessulatum 1986 1986 annelid Glossiphonia complanata Glossiphonia complanata 1986 1986 annelid leeches Erpobdella octoculata 1986 1986 bird Bullfinch Pyrrhula pyrrhula 2016 2017 bird Carrion Crow Corvus corone 2017 2017 bird Chaffinch Fringilla coelebs 2015 2017 bird Chiffchaff Phylloscopus collybita 2014 2016 bird Coot Fulica atra 2014 2014 bird Fieldfare Turdus pilaris 2015 2015 bird Great Tit Parus major 2015 2015 bird Grey Heron Ardea cinerea 2013 2017 bird Jay Garrulus glandarius 1999 1999 bird Kestrel Falco tinnunculus 1999 2015 bird Kingfisher Alcedo atthis 1986 1986 bird Mallard Anas platyrhynchos 2014 2015 bird Marsh Harrier Circus aeruginosus 2000 2000 bird Moorhen Gallinula chloropus 2015 2015 bird Pheasant Phasianus colchicus 2017 2017 bird Robin Erithacus rubecula 2017 2017 bird Spotted Flycatcher Muscicapa striata 1986 1986 bird Tawny Owl Strix aluco 2006 2015 bird Willow Warbler Phylloscopus trochilus 2015 2015 bird Wren Troglodytes troglodytes 2015 2015 bird Yellowhammer Emberiza citrinella 2000 2000 conifer Douglas Fir Pseudotsuga menziesii 2004 2004 conifer European Larch Larix decidua 2004 2004 conifer Lawson's Cypress Chamaecyparis lawsoniana 2004 2004 conifer Scots Pine Pinus sylvestris 1986 2004 crustacean -

December 2019 Suzanne Burgess and Joanna Lindsay
December 2019 Suzanne Burgess and Joanna Lindsay Saving the small things that run the planet Summary Of the 70 species of lacewing recorded in the United Kingdom, at least forty-one of them have been recorded in Scotland, with four only being recorded in Scotland. The Bordered brown lacewing (Megalomus hirtus) was previously known from only two sites in Scotland, at Salisbury’s Crags in Holyrood Park, Edinburgh and at Doonie Point by Bridge of Muchalls in Aberdeenshire. Scottish Natural Heritage (SNH) provided funding to Buglife for year two of the Bordered Brown Lacewing project to run surveys and workshops to raise awareness and improve participant’s identification skills of the different species of lacewing and their allies (alderflies, scorpionflies and snake flies). With the help of volunteers, year two of the project successfully found twenty two adults of the Bordered brown lacewing. Two new areas were discovered at Holyrood Park at rocky outcrops close to St. Anthony’s Chapel and a new population was discovered at Skatie Shore and Perthumie Bay by Stonehaven. A further nine adults were found by Dr Nick Littlewood at six locations from the war memorial south of Stonehaven to Portlethen Village. A total of 264 records of 141 different species of invertebrate, including five species of lacewing, were recorded during surveys and workshops run through this project from six sites: Holyrood Park, in Edinburgh; Hermitage of Braid and Blackford Hill Local Nature Reserve in Edinburgh; Skatie Shore and Perthumie Bay by Stonehaven; Doonie Point by Bridge of Muchalls; St Cyrus National Nature Reserve near Montrose; and Drumpellier Park in North Lanarkshire. -

ABSTRACT the First Through Fifth Instars of the Gypsy Moth Were Tested for Development to Adults on 326 Species of Dicotyledonous Plants in Laboratory Feeding Trials
LABORATORY FEEDING TESTS ON THE DEVELOPMENT OF GYPSY MOTH LARVAE WITH REFERENCE TO PLANT TAXA AND ALLELOCHEMICALS JEFFREY C. MILLER and PAUL E. HANSON DEPARTMENT OF ENTOMOLOGY, OREGON STATE UNIVERSITY, CORVALLIS, OREGON 97331 ABSTRACT The first through fifth instars of the gypsy moth were tested for development to adults on 326 species of dicotyledonous plants in laboratory feeding trials. Among accepted plants, differences in suitability were documented by measuring female pupal weights. The majority of accepted plants belong to the subclasses Dilleniidae, Hamamelidae, and Rosidae. Species of oak, maple, alder, madrone, eucalyptus, poplar, and sumac were highly suitable. Plants belonging to the Asteridae, Caryophyllidae, and Magnoliidae were mostly rejected. Foliage type, new or old, and instar influenced host plant suitability. Larvae of various instars were able to pupate after feeding on foliage of 147 plant species. Of these, 1.01 were accepted by first instars. Larvae from the first through fifth instar failed to molt on foliage of 151 species. Minor feeding occurred on 67 of these species. In general, larvae accepted new foliage on evergreen species more readily than old foliage. The results of these trials were combined with results from three previous studies to provide data on feeding responses of gypsy moth larvae on a total of 658 species, 286 genera, and 106 families of dicots. Allelochemic compositions of these plants were tabulated from available literature and compared with acceptance or rejection by gypsy moth. Plants accepted by gypsy moth generally contain tannins, but lack alkaloids, iridoid monoterpenes, sesquiterpenoids, diterpenoids, and glucosinolates. 2 PREFACE This research was funded through grants from USDA Forest Service cooperative agreement no. -

New Zealand Naturalised Vascular Plant Checklist
NEW ZEALAND NATURALISED VASCULAR PLANT CHECKLIST Clayson Howell; ISBN 0-473-11306-6 John W.D. Sawyer New Zealand Plant Conservation Network November 2006 9 780473 113063 New Zealand naturalised vascular plant checklist November 2006 Clayson J. Howell, John W.D. Sawyer New Zealand Plant Conservation Network P.O. Box 16-102 Wellington New Zealand 6242 E-mail: [email protected] www.nzpcn.org.nz Cover photos (by Jeremy Rolfe): Selaginella kraussiana (Lycophytes), Cestrum elegans (Dicot. trees & shrubs), Cyperus eragrostis (Monocot. herbs: Sedges), Cerastium glomeratum (Dicot. herbs other than composites), Dipogon lignosus (Dicot lianes), Berberis darwinii (Dicot. trees & shrubs), Lonicera japonica (Dicot. lianes), Bomarea caldasii (Monocot. lianes), Pinus radiata (Gymnosperm trees & shrubs), Lilium formosanum (Monocot. herbs other than grasses, orchids, rushes, sedges), Poa annua (Monocot. herbs: Grasses), Clematis vitalba (Dicot. lianes), Adiantum raddianum (Ferns) Main photo: Senecio diaschides (Dicot herbs: Composites). Title page: Asparagus scandens seedling in kauri forest. © Clayson J. Howell, John W.D. Sawyer 2006 ISBN-10: 0-473-12300-2 ISBN-13: 978-0-473-12300-0 Published by: New Zealand Plant Conservation Network P.O. Box 16-102 Wellington 6242 New Zealand E-mail: [email protected] www.nzpcn.org.nz CONTENTS Introduction 1 New Zealand adventive flora – Summary statistics 2 Naturalised plant records in the Flora of New Zealand 2 Naturalised plant checklists in the New Zealand Journal of Botany 2 Species outside Flora or checklists 2 Acknowledgements 4 Bibliography 4 New Zealand naturalised vascular plant checklist – alphabetical 6 iii Cortaderia selloana, one of two species of pampas that are fully naturalised in New Zealand. -

Etude Phytochimique Et Plantes Médicinales Pseudo-Scorodonia
REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE MINISTERE DE L’ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHE SCIENTIFIQUE UNIVERSITE ABOU-BEKR-BELKAID TLEMCEN Faculté des Sciences de la Nature et de la Vie et Sciences de la Terre et de l’Univers Département de Biologie Laboratoire des Produits Naturels « LAPRONA » THÈSE Présentée par Mme BELARBI Karima ép. BOUTERFAS En vue de l’obtention du diplôme de Doctorat en Biologie Cellulaire et Biochimie Option : Produits Naturels, Aspects Nutritionnels et Activités Biologiques Thème Etude phytochimique et activités biologiques de deux plantes médicinales de la région de Tlemcen Teucrium pseudo-Scorodonia Desf. et Erica arborea L. Soutenue le : 12/05/2018 Devant le jury composé de : Président : Mr DJAZIRI Rabeh (Pr) Université de Tlemcen Directrice de thèse : Mme ATIK-BEKKARA Fawzia (Pr) Université de Tlemcen Co-directrice de thèse : Mme BEKHECHI Chahrazed (Pr) Université de Tlemcen Examinateurs : Mr DJEBLI Noureddine (Pr) Université de Mostaganem Mr BELLAHCEN Miloud (Pr) Centre universitaire d’Aïn Témouchent Année universitaire : 2017-2018 Remerciements Avant toute chose, je remercie ALLAH, le tout puissant de m’avoir donné la santé, la patience, la puissance et la volonté pour réaliser ce travail. J’ai eu la chance d’effectuer ce travail de recherche dans le laboratoire des « Produits Naturels » (LAPRONA) du département de Biologie, de la Faculté des Sciences de la Nature et de laVie, des Sciences de la Terre et de l’Univers (SNV/STU), de l’Université Abou Bekr Belkaïd de Tlemcen. Tout d’abord, je souhaite remercier très chaleureusement mon professeur et mon encadreur, Mme ATIK-BEKKARA Fawzia, Professeur à l’Université de Tlemcen et Directrice du laboratoire des Produits Naturels (LAPRONA) qui m’a fait l’honneur de réaliser ce travail sous sa direction. -

A Search for Phylogenetically Informative Pollen Characters in the Subtribe Salviinae (Mentheae: Lamiaceae)
Int. J. Plant Sci. 169(3):455–471. 2008. Ó 2008 by The University of Chicago. All rights reserved. 1058-5893/2008/16903-0011$15.00 DOI: 10.1086/526463 A SEARCH FOR PHYLOGENETICALLY INFORMATIVE POLLEN CHARACTERS IN THE SUBTRIBE SALVIINAE (MENTHEAE: LAMIACEAE) Hye-Kyoung Moon,1,* Stefan Vinckier,y Jay B. Walker,z Erik Smets,*,§ and Suzy Huysmans* *Laboratory of Plant Systematics, Institute of Botany and Microbiology, Katholieke Universiteit Leuven, Kasteelpark Arenberg 31, P.O. Box 2437, BE-3001 Leuven, Belgium; yCenter for Transgene Technology and Gene Therapy, Flanders Institute for Biotechnology (VIB-3), Katholieke Universiteit Leuven, Herestraat 49, BE-3000 Leuven, Belgium; zDepartment of Botany, University of Wisconsin, Madison, Wisconsin 53706, U.S.A.; and §National Herbarium of the Netherlands, Leiden University Branch, P.O. Box 9514, NL-2300 RA Leiden, The Netherlands The pollen morphology and ultrastructure of the subtribe Salviinae were investigated with light, scanning electron, and transmission electron microscopy. In addition, cladistic analyses of the obtained morphological data, supplemented with rbcL data from GenBank, were conducted in order to assess the phylogenetic signal of palynological characters. Salviinae pollen is small to large, oblate to prolate in shape, with a circular to slightly elliptic amb, and mostly hexacolpate. Perovskia abrotanoides appears to be distylous and shows a significant pollen dimorphism between pin and thrum flowers. The sexine ornamentation of the genera Lepechinia and Chaunostoma is perforate, while the ornamentation of other genera is bireticulate. Perovskia expresses a unique type of sexine ornamentation. The sexine ornamentation variation in Salvia has systematic importance but only partly corresponds with current phylogenetic hypotheses. -

Comparative Assessment of Pollen Micromorphology of Salvia Assurgens (Lamiaceae), an Endemic Sage from Mexico
Phytotaxa 458 (3): 183–194 ISSN 1179-3155 (print edition) https://www.mapress.com/j/pt/ PHYTOTAXA Copyright © 2020 Magnolia Press Article ISSN 1179-3163 (online edition) https://doi.org/10.11646/phytotaxa.458.3.1 Comparative assessment of pollen micromorphology of Salvia assurgens (Lamiaceae), an endemic sage from Mexico BRENDA Y. BEDOLLA-GARCÍA1,4, MAYRA CASTRO-MORALES2,5 & CARLOS A. CULTID-MEDINA1,3,6* 1Instituto de Ecología, A. C., Centro Regional del Bajío, Red de Diversidad Biológica del Occidente Mexicano. Apdo. Postal 386, 61600 Pátzcuaro, Michoacán, Mexico. 2Libramiento Ignacio Zaragoza 971 Col. Centro, Código Postal 61600, Pátzcuaro, Michoacán, Mexico. 3CONACyT. Avenida Insurgentes Sur 1582, 03940 Mexico City, Mexico. 4 [email protected]; https://orcid.org/0000-0002-1560-1090 5 [email protected]; https://orcid.org/0000-0001-5726-3010 6 [email protected]; https://orcid.org/0000-0002-4929-8405 *Corresponding author Abstract The present study provides novel information about the pollen of Salvia assurgens. Pollen grains were collected and described based on their observed characters by light microscope and scanning electron microscopy. The species is distinguished from other Mexican salvias by having small pollen grains (14.2 × 18.2 μm), thin primary muri (0.2 μm thick), elongated primary lumina (1.03 μm long) and secondary lumina with relatively few perforations (9, range 5–14). Pollen characters are similar to those of the majority of American salvias. Regarding Mexican sages, there has been little palynological research, and only 23 species of 32 examined have been quantitatively studied. Standardization is needed in different aspects of palynological studies, especially in relation to measurement protocols and data analysis, as well as the increased use of scanning electron microscopy (SEM), since the majority of differences among species are provided by SEM microscope observation.