Neotropical Diptera
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Diptera, Nematocera) from the Mediterranean Region*)
©Zoologische Staatssammlung München;download: http://www.biodiversitylibrary.org/; www.biologiezentrum.at SPIXIANA 14 321-338 München, 3 1 . Oktober 1 991 ISSN0341-8391 New and remarkable species of Mycetophiloidea (Diptera, Nematocera) from the Mediterranean region*) By Norbert Caspers Caspers, N. (1991): New and remarkable species of Mycetophiloidea (Diptera, Nematocera) from the Mediterranean region. - Spixiana 14/3: 321-338. Descriptions of adult males of six neu species of the superfamily Mycetophiloidea (Diptera, Nematocera) from the Mediterranean region are presented: Orfelia persimi- lis, spec. nov. (family Keroplatidae) from Sardinia (Italy); Ectrepesthoneura chand- leri, spec. nov. from Crete (Greece), Leia umbrosa, spec. nov. from Corsica (France), Anatella longiflagellata, spec. nov. and Phronia incisa spec. nov. from Turkey, Sceptonia curvisetosa, spec. nov. from Greece, last five belonging to the family Mycetophilidae sensu Malloch (1917). In addition, new findings of two remarkable and hitherto rarely found species of Mycetophiloidea (Sciopbila kasbmirensis Zaitzev, Phronia tyrrhenica Edwards) are presented. Prof. Dr. Norbert Caspers, St.-Maternus-Eck 14 a, W-5067 Bechen, Germanv Introduction Since the early seventies Dr. H. Malicky (Biological Station Lunz, Austria) has been performing many entomological field trips to Southern Europe, with the aim to clear up the systematics and the biogeographic distribution of the caddis flies (Trichoptera) of that previously rather neglected faunal region. Besides Trichoptera he also collected a comprehensive material of many other aquatic insect groups, which meanwhile has been worked upon and has been published bv different authors. Among the "non-target-groups" of this study the portion of the fungus gnats (Mycetophiloidea) was relatively low as compared to all aquatic insect groups. -

ARTHROPOD COMMUNITIES and PASSERINE DIET: EFFECTS of SHRUB EXPANSION in WESTERN ALASKA by Molly Tankersley Mcdermott, B.A./B.S
Arthropod communities and passerine diet: effects of shrub expansion in Western Alaska Item Type Thesis Authors McDermott, Molly Tankersley Download date 26/09/2021 06:13:39 Link to Item http://hdl.handle.net/11122/7893 ARTHROPOD COMMUNITIES AND PASSERINE DIET: EFFECTS OF SHRUB EXPANSION IN WESTERN ALASKA By Molly Tankersley McDermott, B.A./B.S. A Thesis Submitted in Partial Fulfillment of the Requirements for the Degree of Master of Science in Biological Sciences University of Alaska Fairbanks August 2017 APPROVED: Pat Doak, Committee Chair Greg Breed, Committee Member Colleen Handel, Committee Member Christa Mulder, Committee Member Kris Hundertmark, Chair Department o f Biology and Wildlife Paul Layer, Dean College o f Natural Science and Mathematics Michael Castellini, Dean of the Graduate School ABSTRACT Across the Arctic, taller woody shrubs, particularly willow (Salix spp.), birch (Betula spp.), and alder (Alnus spp.), have been expanding rapidly onto tundra. Changes in vegetation structure can alter the physical habitat structure, thermal environment, and food available to arthropods, which play an important role in the structure and functioning of Arctic ecosystems. Not only do they provide key ecosystem services such as pollination and nutrient cycling, they are an essential food source for migratory birds. In this study I examined the relationships between the abundance, diversity, and community composition of arthropods and the height and cover of several shrub species across a tundra-shrub gradient in northwestern Alaska. To characterize nestling diet of common passerines that occupy this gradient, I used next-generation sequencing of fecal matter. Willow cover was strongly and consistently associated with abundance and biomass of arthropods and significant shifts in arthropod community composition and diversity. -
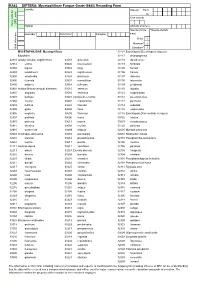
Ra82 Diptera
RA82 DIPTERA: Mycetophilinae Fungus Gnats (6480) Recording Form Locality Date(s) from: to: Vice county GPS users Grey cells for Habitat Altitude (metres) Source (circle *Source details Recorder Determiner Compiler one) Field 1 Museum* 2 Grid reference Literature* 3 MYCETOPHILIDAE: Mycetophilinae 33101 Exechiopsis (Exechiopsis) clypeata Exechiini 33117 dryaspagensis 32801 Allodia (Allodia) anglofennica 33519 griseolum33118 dumitrescae 32912 embla 33526 intermedium33119 fimbriata 32802 lugens 33522 kingi33120 furcata 32803 lundstroemi 33523 nigrofuscum33106 hammi 32804 ornaticollis 33524 proximum33107 indecisa 32806 truncata 33527 rosmellitum33108 intersecta 32805 zaitzevi 33514 ruficorne33109 jenkinsoni 32901 Allodia (Brachycampta) alternans 33515 serenum33110 ligulata 32910 angulata 33516 sericoma33121 magnicauda 32903 barbata 33601 Cordyla brevicornis33112 pseudindecisa 32904 czernyi 33602 crassicornis33113 pulchella 32909 foliifera 33603 fasciata33114 subulata 32905 grata 33604 fissa33115 unguiculata 32906 neglecta 33605 flaviceps33116 Exechiopsis (Xenexechia) crucigera 32907 pistillata 33606 fusca33002 leptura 32915 protenta 33613 insons33003 membranacea 32914 silvatica 33608 murina33122 pollicata 32916 westerholti 33609 nitidula32605 Myrosia maculosa 32602 Allodiopsis domestica 33610 parvipalpis32601 Notolopha cristata 32610 korolevi 33614 pseudomurina32701 Pseudexechia aurivernica 32607 rustica 33611 pusilla32706 monica 32212 Anatella alpina 33612 semiflava32705 parallela 32213 ankeli 33201 Exechia bicincta32703 trisignata 32216 bremia -

Recent Noteworthy Findings of Fungus Gnats from Finland and Northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae)
Biodiversity Data Journal 2: e1068 doi: 10.3897/BDJ.2.e1068 Taxonomic paper Recent noteworthy findings of fungus gnats from Finland and northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae) Jevgeni Jakovlev†, Jukka Salmela ‡,§, Alexei Polevoi|, Jouni Penttinen ¶, Noora-Annukka Vartija# † Finnish Environment Insitutute, Helsinki, Finland ‡ Metsähallitus (Natural Heritage Services), Rovaniemi, Finland § Zoological Museum, University of Turku, Turku, Finland | Forest Research Institute KarRC RAS, Petrozavodsk, Russia ¶ Metsähallitus (Natural Heritage Services), Jyväskylä, Finland # Toivakka, Myllyntie, Finland Corresponding author: Jukka Salmela ([email protected]) Academic editor: Vladimir Blagoderov Received: 10 Feb 2014 | Accepted: 01 Apr 2014 | Published: 02 Apr 2014 Citation: Jakovlev J, Salmela J, Polevoi A, Penttinen J, Vartija N (2014) Recent noteworthy findings of fungus gnats from Finland and northwestern Russia (Diptera: Ditomyiidae, Keroplatidae, Bolitophilidae and Mycetophilidae). Biodiversity Data Journal 2: e1068. doi: 10.3897/BDJ.2.e1068 Abstract New faunistic data on fungus gnats (Diptera: Sciaroidea excluding Sciaridae) from Finland and NW Russia (Karelia and Murmansk Region) are presented. A total of 64 and 34 species are reported for the first time form Finland and Russian Karelia, respectively. Nine of the species are also new for the European fauna: Mycomya shewelli Väisänen, 1984,M. thula Väisänen, 1984, Acnemia trifida Zaitzev, 1982, Coelosia gracilis Johannsen, 1912, Orfelia krivosheinae Zaitzev, 1994, Mycetophila biformis Maximova, 2002, M. monstera Maximova, 2002, M. uschaica Subbotina & Maximova, 2011 and Trichonta palustris Maximova, 2002. Keywords Sciaroidea, Fennoscandia, faunistics © Jakovlev J et al. This is an open access article distributed under the terms of the Creative Commons Attribution License (CC BY 4.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited. -

Of Greece, Its Islands
CHANDLERet al.: 255-314 - Studia dipterologica 12 (2005) Heft 2 ISSN 0945-3954 The Fungus Gnats (Diptera: Bolitophilidae, Diadocidiidae, Ditomyiidae , Keroplatidae and Mycetophilidae) of Greece, its islands and Cyprus [Die Pilzmiicken (Diptera: Bolitophilidae, Diadocidiidae, Ditomyiidae, Keroplatidae und Mycetophilidae) Griechenlands und seiner Inseln sowie Zypern4 1 by Peter J. CHANDLER, Dimitar N. BECHEV and Norbert CASPERS Mclksham (UK) Plovdiv (Bulgaria) Bechen (Gernlany) - - -. - ~ Abstract The spccics of fungu\ gnats (Bolitophilidae, Diadoc~dildae,Ditomyiidac. Keroplat~d:~eand Mycetophilidae) o~urringin Greece and Cyprus are reviewed. Altogether 201 species :Ire recorded, 189 for Greece and 69 for Cyprus. Of these 126 specie5 arc newly recorded fol. Greece and 36 arc newly recorded for Cyprus. The following new taxa arc described from Greece: Macrorrhyrtcha ibis spec. nov., M. pelargos spec. nov., M. laconica spec. nov., Macrocera critica spec. nov., Docosia cephaloniae spec. nov., D. enos spec. nov., D. pa- siphae spec. nov., Megophthalmidia illyrica spec. nov.. M. ionica spec. nov., M. pytho spec. nov., Mycomya thrakis spec. nov., Allocolocera scheria spec. nov., Sciophila pandora spec. nov., Ryrnosia labyrinthos spec. nov.; M. ill\,ric,cr is also recorded troln Croc~lia.The follow- ing ncw taa are described from Cyprus: Macrocera cypriaca spec. nov., Megophthalmidia alrzicola spec. nov., M. cedricola spec. nov. The following neu synonymies are propod: M!,c,c~r~iwrenuis I WXLKER,1856) = M. interniissa PL.ASSMA~N,l984 syn. nov., Plrror~rtr~1.illi.s- torri DLIFI>ZICKI,1889 = P rnciscr CASFERS,1991 syn. nov. A key is provided for thc western Palaearctic specie5 of M(ic-i.orrh~~~ic-IrciWI~~ERTZ. -

Zootaxa, the Fungus Gnats (Diptera: Bolitophilidae
Zootaxa 2318: 450–506 (2009) ISSN 1175-5326 (print edition) www.mapress.com/zootaxa/ Article ZOOTAXA Copyright © 2009 · Magnolia Press ISSN 1175-5334 (online edition) The fungus gnats (Diptera: Bolitophilidae, Keroplatidae, Mycetophilidae) of Sardinia, with description of six new species* PETER J. CHANDLER 606B Berryfield Lane, Melksham, Wilts SN12 6EL, United Kingdom. E-mail: [email protected] *In: Cerretti, P., Mason, F., Minelli, A., Nardi, G. & Whitmore, D. (Eds), Research on the Terrestrial Arthropods of Sardinia (Italy). Zootaxa, 2318, 1–602. Table of contents Abstract . .450 Introduction . .451 Study area . .452 Material and methods . .452 Abbreviations . .453 Sampling sites . .454 Faunistic list . .456 Bolitophilidae . .456 Keroplatidae, Keroplatinae, Keroplatini . .456 Orfeliini . .457 Macrocerinae . .462 Mycetophilidae, Gnoristinae . .465 Leiinae . .469 Mycetophilinae, Exechiini . .472 Mycetophilini . .480 Mycomyinae . .492 Sciophilinae . .495 Discussion . .500 Acknowledgements . .501 References . .502 Abstract The fungus gnat fauna of Sardinia is reviewed and data presented for all species recorded. Altogether one species of Bolitophilidae, 16 species of Keroplatidae and 105 species of Mycetophilidae are recognised as occurring in Sardinia. As the bolitophilid and two of the mycetophilid species are represented only by females and are not determined to species level, the total confirmed Sardinian list stands at 119 species. Four species of Keroplatidae and 19 species of Mycetophilidae are new to the total Italian fauna, whereas three species of Keroplatidae and 32 species of Mycetophilidae are newly recorded for the island of Sardinia. Six species are described as new to science: two Keroplatidae (Urytalpa juliae sp. nov., Macrocera nuragica sp. nov.) and four Mycetophilidae (Boletina ichnusa sp. nov., Trichonta sandalyon sp. -

Volume 2, Chapter 12-19: Terrestrial Insects: Holometabola-Diptera
Glime, J. M. 2017. Terrestrial Insects: Holometabola – Diptera Nematocera 2. In: Glime, J. M. Bryophyte Ecology. Volume 2. 12-19-1 Interactions. Ebook sponsored by Michigan Technological University and the International Association of Bryologists. eBook last updated 19 July 2020 and available at <http://digitalcommons.mtu.edu/bryophyte-ecology2/>. CHAPTER 12-19 TERRESTRIAL INSECTS: HOLOMETABOLA – DIPTERA NEMATOCERA 2 TABLE OF CONTENTS Cecidomyiidae – Gall Midges ........................................................................................................................ 12-19-2 Mycetophilidae – Fungus Gnats ..................................................................................................................... 12-19-3 Sciaridae – Dark-winged Fungus Gnats ......................................................................................................... 12-19-4 Ceratopogonidae – Biting Midges .................................................................................................................. 12-19-6 Chironomidae – Midges ................................................................................................................................. 12-19-9 Belgica .................................................................................................................................................. 12-19-14 Culicidae – Mosquitoes ................................................................................................................................ 12-19-15 Simuliidae – Blackflies -

ARTHROPODA Subphylum Hexapoda Protura, Springtails, Diplura, and Insects
NINE Phylum ARTHROPODA SUBPHYLUM HEXAPODA Protura, springtails, Diplura, and insects ROD P. MACFARLANE, PETER A. MADDISON, IAN G. ANDREW, JOCELYN A. BERRY, PETER M. JOHNS, ROBERT J. B. HOARE, MARIE-CLAUDE LARIVIÈRE, PENELOPE GREENSLADE, ROSA C. HENDERSON, COURTenaY N. SMITHERS, RicarDO L. PALMA, JOHN B. WARD, ROBERT L. C. PILGRIM, DaVID R. TOWNS, IAN McLELLAN, DAVID A. J. TEULON, TERRY R. HITCHINGS, VICTOR F. EASTOP, NICHOLAS A. MARTIN, MURRAY J. FLETCHER, MARLON A. W. STUFKENS, PAMELA J. DALE, Daniel BURCKHARDT, THOMAS R. BUCKLEY, STEVEN A. TREWICK defining feature of the Hexapoda, as the name suggests, is six legs. Also, the body comprises a head, thorax, and abdomen. The number A of abdominal segments varies, however; there are only six in the Collembola (springtails), 9–12 in the Protura, and 10 in the Diplura, whereas in all other hexapods there are strictly 11. Insects are now regarded as comprising only those hexapods with 11 abdominal segments. Whereas crustaceans are the dominant group of arthropods in the sea, hexapods prevail on land, in numbers and biomass. Altogether, the Hexapoda constitutes the most diverse group of animals – the estimated number of described species worldwide is just over 900,000, with the beetles (order Coleoptera) comprising more than a third of these. Today, the Hexapoda is considered to contain four classes – the Insecta, and the Protura, Collembola, and Diplura. The latter three classes were formerly allied with the insect orders Archaeognatha (jumping bristletails) and Thysanura (silverfish) as the insect subclass Apterygota (‘wingless’). The Apterygota is now regarded as an artificial assemblage (Bitsch & Bitsch 2000). -

Frontenac Provincial Park
FRONTENAC PROVINCIAL PARK One Malaise trap was deployed at Frontenac Provincial Park in 2014 (44.51783, -76.53944, 176m ASL; Figure 1). This trap collected arthropods for twenty weeks from May 9 – September 25, 2014. All 10 Malaise trap samples were processed; every other sample was analyzed using the individual specimen protocol while the second half was analyzed via bulk analysis. A total of 3372 BINs were obtained. Half of the BINs captured were flies (Diptera), followed by bees, ants and wasps (Hymenoptera), moths and butterflies (Lepidoptera), and true bugs (Hemiptera; Figure 2). In total, 750 arthropod species were named, representing 24.6% of the BINs from the site (Appendix 1). All but 1 of the BINs were assigned Figure 1. Malaise trap deployed at Frontenac at least to family, and 58.2% were assigned to a genus Provincial Park in 2014. (Appendix 2). Specimens collected from Frontenac represent 232 different families and 838 genera. Diptera Hymenoptera Lepidoptera Hemiptera Coleoptera Trombidiformes Psocodea Trichoptera Araneae Mesostigmata Entomobryomorpha Thysanoptera Neuroptera Orthoptera Sarcoptiformes Blattodea Mecoptera Odonata Symphypleona Ephemeroptera Julida Opiliones Figure 2. Taxonomy breakdown of BINs captured in the Malaise trap at Frontenac. APPENDIX 1. TAXONOMY REPORT Class Order Family Genus Species Arachnida Araneae Clubionidae Clubiona Clubiona obesa Dictynidae Emblyna Emblyna annulipes Emblyna sublata Gnaphosidae Cesonia Cesonia bilineata Linyphiidae Ceraticelus Ceraticelus fissiceps Pityohyphantes Pityohyphantes -

Kjaerandsen Sciaroidea WIP.Pdf
Species recognition trade-off between structural wing colours and terminalia in fungus gnats ? J. Kjaerandsen Museum of Zoology Lund University Sweden Structural colours in flies Reflective scales in Diptera – Mosquitoes: Toxorhynchites manicatus (Japan) Reflective body scales in fungus gnats – only in the genus Allactoneura ? Hymenoptera: Eulophidae PhD student Ekaterina Shevtsova Wings imbedded in a medium or studied on a white background will not display their structural colours Slide with wings embedded in Canada balsam Dry specimens studied on a pure white background Mycetophilidae: Rymosia fasciata Keroplatidae: Proceroplatus scalprifera WIPs — Wing Interference Patterns i for interference Bolitophila occlusa Hybotidae: Ocydromia glabricula Cordyla sp. (California) Exechia nugatoria Photo: “Klaas” at Diptera.info, 2008 (= nigroscutellata) (California) Photos: Peter Kerr, 2008 My photo of the same species’ WIP Photos of structural wing colours on internet WIPs — Wing Interference Patterns i for interference • — Genetics of pigment patterns • — Thin Film Interference • — Newton Scale Metering • — Exechiini • — Lygistorrhinidae • — Keroplatidae • — The trade-off Pigmentation in brown, yellow and black: Spatiotemporally regulated by yellow and ebony MELANINS Leia Proceroplatus (Japan) (New Caledonia) Scientists unlock mystery of animal colour patterns Genetic April 22 control of pigment 2010 patterns T. Werner, S. Koshikawa, T. M. Williams, S. B. Carroll, Nature 464, 1143 (2010) Pigments are only a part of the ”mystery of wing colour -

Smithsonian Miscellaneous Collections Volume 117, Number 3
SMITHSONIAN MISCELLANEOUS COLLECTIONS VOLUME 117, NUMBER 3 RELATIONSHIPS OF CERTAIN GENERA OF FUNGUS GNATS OF THE FAMILY MYCETOPHILIDAE BY F. R. SHAW AND M. M. SHAW Amherst, Mass. (Publication 4053) CITY OF WASHINGTON PUBLISHED BY THE SMITHSONIAN INSTITUTION DECEMBER 27, 1951 Z^t £or5 (g»af(tmore (pv&ee BALTIMOEB, MD., 0. B. A. RELATIONSHIPS OF CERTAIN GENERA OF FUNGUS GNATS OF THE FAMILY MYCETOPHILIDAE By F. R. SHAW and M. M. SHAW i Amherst, Mass. The present study represents a continuation of a preliminary in- vestigation of the possible value of thoracic sclerites in determining the relationships of certain insects. Dr. G. C. Crampton was the first to demonstrate the use of these sclerites as a means of determin- ing the systematic position of insects. In 1925, he published a clas- sical study of the comparative morphology of the thorax of nontipu- loid Nematocera. In 1948 Shaw presented a paper in which he indi- cated the value of thoracic sclerites as an aid in determining the phy- logeny of the Mycetophilidae. Although the number of genera he studied was admittedly small, principles were developed that have been of value in distinguishing the phylogenetic relationships of cer- tain genera. Edwards (1925) was the first to indicate that the structure of thoracic sclerites might be of value in determining generic characters in this group. In his monograph of the British fungus gnats he noted that in certain genera the sclerites differed in form and that such dif- ferences might be of value in separating groups of these insects. 1 We wish to express our thanks to the Society of Sigma Xi for a grant-in- aid that made possible the preparation of the illustrations for this paper. -

American Museum Novitates
AMERICAN MUSEUM NOVITATES Number 3915, 19 pp. December 18, 2018 A new species of Eumanota Edwards (Diptera: Mycetophilidae: Manotine) from Colombia: evidence for a pseudogondwanan pattern DALTON S. AMORIM,1 SARAH S. OLIVEIRA,2 AND ANDREA C. HENAO-SEPÚLVEDA3 ABSTRACT Eumanota wolffae, sp. nov., is described from the high Andean forests of Colombia. This is the first Neotropical species of the clade of non-Manota genera of the mycetophilid subfamily Manotinae—to date known entirely from the Oriental and the northwestern Australasian regions, and in Baltic amber. With the other species of Eumanota Edwards, this species shares, among other features, a largely developed third maxillary palpomere, projecting beyond the base of fourth palpomere, with a wide sensorial pit, a flat inner face, and the last palpomere at least 4× the length of the fourth palpomere. The systematic position of the Colombian species within the Manotinae is addressed. The biogeographic significance of Eumanota in the Neo- tropical region is discussed, interpreted here as a concurrent incidence of a circumtropical pattern and Gondwanan distributions, in other words, a particular pattern of biogeographic pseudocongruence, referred to here as a “pseudogondwanan pattern.” This pattern is associated with an early Cenozoic tropical biota over Laurasian terranes that expanded its distribution to the south (in the Americas, Africa, and Australasia), followed by large-scale extinction of Nearctic and Palearctic representatives due to global cooling in the Neogene. A discussion is provided about the correlation between these patterns and evidence of Late Cretaceous-Paleo- gene tropical floras over Laurasian terranes. 1 Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, Universidade de São Paulo, São Paulo, Brazil.