Freshwater Animal Diversity Assessment Developments in Hydrobiology 198
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Amphidromy in Shrimps: a Life Cycle Between Rivers and the Sea
Lat. Am. J. Aquat. Res., 41(4): 633-650, 2013 Amphidromy in shrimps: a life cycle 633 “Studies on Freshwater Decapods in Latin America” Ingo S. Wehrtmann & Raymond T. Bauer (Guest Editors) DOI: 103856/vol41-issue4-fulltext-2 Review Amphidromy in shrimps: a life cycle between rivers and the sea Raymond T. Bauer1 1Department of Biology, University of Louisiana at Lafayette, Lafayette, Louisiana, 70504-2451 USA ABSTRACT. Amphidromy is a diadromous life history pattern, common in tropical and subtropical freshwater caridean shrimps, in which adults live, breed and spawn small-sized embryos in freshwater but have extended larval development (ELD) in marine waters. Most completely freshwater species spawn large embryos with either direct or abbreviated larval development (ALD). An important benefit of amphidromy is dispersal among river systems via marine larvae, which increases their access to alternative habitats. Thus, amphidromous species have much broader geographic distributions than closely related completely freshwater ones with ALD. ALD and freshwater ELD species appear to have evolved from amphidromous species with marine ancestors. Delivery of larvae to the sea in many amphidromous species is accomplished by upstream hatching and river drift of larvae to the sea. In other species, the females themselves apparently migrate down to marine waters to spawn. After development, the postlarvae must find a river mouth and migrate upstream to the adult habitat. Migrations occur at night, with juveniles swimming or crawling along the river or stream bank. Larvae are released during the wet or flood season of the year, while juvenile migrations take place during the dry or low-flow season. -

Ophthalmotilapia Boops (BOULENGER, 1901)
DCG_Info_05_2016_HR_20160420_Karlsson_ohne_Haustein_DCG_Info 20.04.2016 06:36 Seite 106 Ophthalmotilapia boops (BOULENGER, 1901) Wolfgang Staeck Abb. 1. Männchen von Ophthalmotilapia boops über seiner Laichgrube auf einem Felsen bei Kampemba Point (Kleines Bild: Ophthalmotilapia boops und Tropheus bei Kampemba Point) Meine erste Begegnung mit diesem un- Beide sprachen kein Englisch, sondern geografische Farbvariante von Oph- gewöhnlichen Tanganjikasee-Bunt- unterhielten sich gewöhnlich auf Sua- thalmotilapia ventralis hielt, zu fangen, barsch liegt nun bereits fast vierzig heli, das ich nicht beherrschte. Da in weil wir nicht über die dazu benötigte Jahre zurück. Im Jahre 1977 bereiste Burundi aber neben der afrikanischen Ausrüstung verfügten, um sie heil bis ich zusammen mit zwei Fängern des Muttersprache auch Französisch ge- in das weit entfernte Kigoma zu trans- damals in Dar es Salaam lebenden Ex- sprochen wurde, war die Kommunika- portieren. porteurs Misha Fainzilber in einem der tion in unserer kleinen Reisegruppe dort üblichen motorisierten Fischer- kein unlösbares Problem. Systematik und Merkmale von Oph- boote von Kigoma aus die zu Tansania thalmotilapia gehörende Ostküste des Tanganjikasees Ich erinnere mich noch genau über in südlicher Richtung bis nach Kipili. meine Begeisterung, als ich in der Um- Ophthalmotilapia boops gehört zu den Ziel unserer fast drei Wochen dauern- gebung der Ortschaft Kipili schnor- ersten Arten, die aus dem Tanganjika- den Entdeckungsreise war es, für den chelte und dort im Uferbereich der see beschrieben wurden, denn die bei- Exporteur eine erste Bestandsaufnahme Insel Kerenge erstmals die auffallend den Typusexemplare sind von dem der in diesem Gebiet vorkommenden kontrastreich tief schwarz und leuch- englischen Forschungsreisenden MOORE Buntbarsche vorzunehmen. tend hellblau gezeichneten Männchen bereits gegen Ende des 19. -

Decapod Crustacean Grooming: Functional Morphology, Adaptive Value, and Phylogenetic Significance
Decapod crustacean grooming: Functional morphology, adaptive value, and phylogenetic significance N RAYMOND T.BAUER Center for Crustacean Research, University of Southwestern Louisiana, USA ABSTRACT Grooming behavior is well developed in many decapod crustaceans. Antennular grooming by the third maxillipedes is found throughout the Decapoda. Gill cleaning mechanisms are qaite variable: chelipede brushes, setiferous epipods, epipod-setobranch systems. However, microstructure of gill cleaning setae, which are equipped with digitate scale setules, is quite conservative. General body grooming, performed by serrate setal brushes on chelipedes and/or posterior pereiopods, is best developed in decapods at a natant grade of body morphology. Brachyuran crabs exhibit less body grooming and virtually no specialized body grooming structures. It is hypothesized that the fouling pressures for body grooming are more severe in natant than in replant decapods. Epizoic fouling, particularly microbial fouling, and sediment fouling have been shown r I m ans of amputation experiments to produce severe effects on olfactory hairs, gills, and i.icubated embryos within short lime periods. Grooming has been strongly suggested as an important factor in the coevolution of a rhizocephalan parasite and its anomuran host. The behavioral organization of grooming is poorly studied; the nature of stimuli promoting grooming is not understood. Grooming characters may contribute to an understanding of certain aspects of decapod phylogeny. The occurrence of specialized antennal grooming brushes in the Stenopodidea, Caridea, and Dendrobranchiata is probably not due to convergence; alternative hypotheses are proposed to explain the distribution of this grooming character. Gill cleaning and general body grooming characters support a thalassinidean origin of the Anomura; the hypothesis of brachyuran monophyly is supported by the conservative and unique gill-cleaning method of the group. -

W7192e19.Pdf
click for previous page 952 Shrimps and Prawns Sicyoniidae SICYONIIDAE Rock shrimps iagnostic characters: Body generally Drobust, with shell very hard, of “stony” grooves appearance; abdomen often with deep grooves and numerous tubercles. Rostrum well developed and extending beyond eyes, always bearing more than 3 upper teeth (in- cluding those on carapace); base of eyestalk with styliform projection on inner surface, but without tubercle on inner border. Both upper and lower antennular flagella of similar length, attached to tip of antennular peduncle. 1 Carapace lacks both postorbital and postantennal spines, cervical groove in- distinct or absent. Exopod present only on first maxilliped. All 5 pairs of legs well devel- 2 oped, fourth leg bearing a single well-devel- 3rd and 4th pleopods 4 single-branched oped arthrobranch (hidden beneath 3 carapace). In males, endopod of second pair 5 of pleopods (abdominal appendages) with appendix masculina only. Third and fourth pleopods single-branched. Telson generally armed with a pair of fixed lateral spines. Colour: body colour varies from dark brown to reddish; often with distinct spots or colour markings on carapace and/or abdomen - such colour markings are specific and very useful in distinguishing the species. Habitat, biology, and fisheries: All members of this family are marine and can be found from shallow to deep waters (to depths of more than 400 m). They are all benthic and occur on both soft and hard bottoms. Their sizes are generally small, about 2 to 8 cm, but some species can reach a body length over 15 cm. The sexes are easily distinguished by the presence of a large copulatory organ (petasma) on the first pair of pleopods of males, while the females have the posterior thoracic sternites modified into a large sperm receptacle process (thelycum) which holds the spermatophores or sperm sacs (usually whitish or yellowish in colour) after mating. -

Gyraulus Laevis in Nederland 87
Kuijper: Gyraulus laevis in Nederland 87 Gyraulus laevis (Mollusca: Planorbidae) in Nederland door W.J. Kuijper Enkele recente waarnemingen van een van onze zeldzaamste planorbiden waren de aanleiding tot het samenstellen van een over- laevis zicht van alle van Nederland bekende vondsten van Gyraulus (Alder, 1838). Tot voor kort waren slechts enkele vindplaatsen van dit dier bekend (Janssen & De Vogel, 1965: 75). Hierbij komt het betrof feit dat het in een aantal gevallen slechts een enkel exemplaar en de soort niet meer teruggevonden kon worden. Het volgende geeft een chronologisch overzicht van de waarnemingen van Gyraulus laevis in Nederland. Voor zover beschikbaar zijn diverse gegevens van de vindplaatsen vermeld. WAARNEMINGEN 1. Koudekerke (bij Middelburg), buitenplaats Vijvervreugd, ± 1890. Fraai vers materiaal: 69 exemplaren in de collectie Schep- de man (Zoölogisch Museum, Amsterdam). Later niet meer vermeld, buitenplaats bestaat niet meer (Kuiper, 1944: 6). Dit materiaalwerd verzameld door Dr. IJ. Keijzer en is vermeld in: Verslag over 1885-1893 van het Zeeuwsch Genootschap der Wetenschappen, blz. 88 (volgens Mevr. Dr. W.S.S. van der Feen - van Benthem Jut- ting). Verder is materiaal van deze vindplaats in de collecties van het Zeeuwsch Museum te Middelburg (2 ex.) en het Natuurhistorisch Museum te Enschede (6 ex.) aanwezig. 2. Warmond (bij Leiden), in de Leede, januari 1916 (Van Ben- them Jutting, 1933: 179). Dit materiaal werd verzameld door P.P. de Koning; in het Rijksmuseum van Natuurlijke Historie te Leiden bevinden zich vier schelpen van deze vindplaats, waarvan twee een doorsnede van ca. 4 mm bereiken. 3. 't Zand (bij Roodeschool), Oostpolder, 1927 (Van Benthem Jutting, 1947: 66). -

Genus Physefocaris, Gen. Nov. Carapace Greatly Inflated
196 Zoologica: Neiv York Zoological Society I" XXV: 11 300 #00 £00 600 700 S00 900 /000 Text-figure 61. Variation in the average carapace length of catches of Para- pandalus richardi made at 100 fathom intervals. Family Physetocaridae, fam. nov. Rostrum present as a broad prolongation of the carapace. First pereio- pods simple. Second pereiopods chelate, with the carpus segmented. No exopods on the third maxillipeds or any of the pereiopods. Terminal joint of the second maxillipeds not applied as a strip to the end of the preceding joint. Mandible without an incisor process or palp. Genus Physefocaris, gen. nov. Carapace greatly inflated. Carpus of second pereiopods consisting of four segments; chela flattened with a very short, broad dactyl. Branchial formula as follows: VII VIII IX x XI XII XIII XIV Podobranchiae ep. ep. ep. ep. ep. ep. Arthrobranchiae Pleurobranchi ae 1 1 1 1 1 Physetocaris microphthalma, sp. nov. Text-figs. 62 and 63. Types: Holotype female (?) Cat. No. 30,523, Department of Tropical Research, New York Zoological Society; Net 798; July 15, 1930; 600 fathoms. 1940] Chace: Bathypelagic, Caridean Crustacea- 197 Physetocaris microphthalma. Holotype. X 6.00. One female (?) ; Net 983, 990, 997, 1003, 1014, 1015, 1016, 1102, 1108, 1115, 1121, 1131, 1137 (?), 1138 (?), 1144, 1149 or 1150; June 2 to August 8, 1931; 500 fathoms. Diagnosis: Carapace with two lateral carinae. Abdomen without any dorsal carinae or spines. Telson deeply sulcate dorsally and broadly truncate at the tip. Eyes very small and set on outside of stalks. Description: Integument extremely thin and fragile. Carapace markedly inflated dorsally and anteriorly to form a very broad, inflated rostrum. -

And Giant Guitarfish (Rhynchobatus Djiddensis)
VIRAL DISCOVERY IN BLUEGILL SUNFISH (LEPOMIS MACROCHIRUS) AND GIANT GUITARFISH (RHYNCHOBATUS DJIDDENSIS) BY HISTOPATHOLOGY EVALUATION, METAGENOMIC ANALYSIS AND NEXT GENERATION SEQUENCING by JENNIFER ANNE DILL (Under the Direction of Alvin Camus) ABSTRACT The rapid growth of aquaculture production and international trade in live fish has led to the emergence of many new diseases. The introduction of novel disease agents can result in significant economic losses, as well as threats to vulnerable wild fish populations. Losses are often exacerbated by a lack of agent identification, delay in the development of diagnostic tools and poor knowledge of host range and susceptibility. Examples in bluegill sunfish (Lepomis macrochirus) and the giant guitarfish (Rhynchobatus djiddensis) will be discussed here. Bluegill are popular freshwater game fish, native to eastern North America, living in shallow lakes, ponds, and slow moving waterways. Bluegill experiencing epizootics of proliferative lip and skin lesions, characterized by epidermal hyperplasia, papillomas, and rarely squamous cell carcinoma, were investigated in two isolated poopulations. Next generation genomic sequencing revealed partial DNA sequences of an endogenous retrovirus and the entire circular genome of a novel hepadnavirus. Giant Guitarfish, a rajiform elasmobranch listed as ‘vulnerable’ on the IUCN Red List, are found in the tropical Western Indian Ocean. Proliferative skin lesions were observed on the ventrum and caudal fin of a juvenile male quarantined at a public aquarium following international shipment. Histologically, lesions consisted of papillomatous epidermal hyperplasia with myriad large, amphophilic, intranuclear inclusions. Deep sequencing and metagenomic analysis produced the complete genomes of two novel DNA viruses, a typical polyomavirus and a second unclassified virus with a 20 kb genome tentatively named Colossomavirus. -
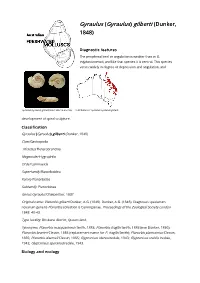
Gyraulus) Gilberti (Dunker, 1848
Gyraulus (Gyraulus) gilberti (Dunker, 1848) Diagnostic features The peripheral keel or angulation is weaker than in G. edgbastonensis, and like that species it is central. This species varies widely in degree of depression and angulation, and Gyraulus (Gyraulus) gilberti (adult size 4.8-5.5 mm) Distribution of Gyraulus (Gyraulus) gilberti. development of spiral sculpture. Classification Gyraulus (Gyraulus) gilberti (Dunker, 1848) Class Gastropoda I nfraclass Heterobranchia Megaorder Hygrophila Order Lymnaeida Superfamily Planorboidea Family Planorbidae Subfamily: Planorbinae Genus Gyraulus Charpentier, 1837 Original name: Planorbis gilberti Dunker, A.G. (1848). Dunker, A.G. (1848). Diagnoses specierum novarum generis Planorbis collection is Cumingianae. Proceedings of the Zoological Society London 1848: 40-43. Type locality: Brisbane district, Queensland. Synonyms: Planorbis macquariensis Smith, 1883; Planorbis fragilis Smith, 1883 (non Dunker, 1850); Planorbis brazieri Clessin, 1885 (replacement name for P. fragilis Smith); Planorbis planissimus Clessin, 1885; Planorbis daemeli Clessin, 1885; Glyptanisus idenusredale, 1943; Glyptanisus stabilis redale, 1943; Glyptanisus speranusredale, 1943. Biology and ecology This species lives in water weeds and other vegetation in ponds, billabongs, swamps and sluggish streams and rivers in tropical and subtropical eastern Australia. Feeds on detritus. Egg mass presumably a jelly strip containing small eggs. Development direct. Brown (2001) described the anatomy of this species. This species is an intermediate host for the stomach fluke Orthocoelium streptocoelium (Boray, 1982; Beesley et al., 1998). Distribution This species occurs throughout eastern Australia, from Cape York to northern New South Wales. Notes G. isingi and/or G.waterhousei may possibly be conspecific with this species (Brown, 2001). Further reading Beesley, P. L., Ross, G. J. -

Anisus Vorticulus (Troschel 1834) (Gastropoda: Planorbidae) in Northeast Germany
JOURNAL OF CONCHOLOGY (2013), VOL.41, NO.3 389 SOME ECOLOGICAL PECULIARITIES OF ANISUS VORTICULUS (TROSCHEL 1834) (GASTROPODA: PLANORBIDAE) IN NORTHEAST GERMANY MICHAEL L. ZETTLER Leibniz Institute for Baltic Sea Research Warnemünde, Seestr. 15, D-18119 Rostock, Germany Abstract During the EU Habitats Directive monitoring between 2008 and 2010 the ecological requirements of the gastropod species Anisus vorticulus (Troschel 1834) were investigated in 24 different waterbodies of northeast Germany. 117 sampling units were analyzed quantitatively. 45 of these units contained living individuals of the target species in abundances between 4 and 616 individuals m-2. More than 25.300 living individuals of accompanying freshwater mollusc species and about 9.400 empty shells were counted and determined to the species level. Altogether 47 species were identified. The benefit of enhanced knowledge on the ecological requirements was gained due to the wide range and high number of sampled habitats with both obviously convenient and inconvenient living conditions for A. vorticulus. In northeast Germany the amphibian zones of sheltered mesotrophic lake shores, swampy (lime) fens and peat holes which are sun exposed and have populations of any Chara species belong to the optimal, continuously and densely colonized biotopes. The cluster analysis emphasized that A. vorticulus was associated with a typical species composition, which can be named as “Anisus-vorticulus-community”. In compliance with that both the frequency of combined occurrence of species and their similarity in relative abundance are important. The following species belong to the “Anisus-vorticulus-community” in northeast Germany: Pisidium obtusale, Pisidium milium, Pisidium pseudosphaerium, Bithynia leachii, Stagnicola palustris, Valvata cristata, Bathyomphalus contortus, Bithynia tentaculata, Anisus vortex, Hippeutis complanatus, Gyraulus crista, Physa fontinalis, Segmentina nitida and Anisus vorticulus. -

Note Food and Feeding Habits of Macrobrachium
Indian J. Fish., 60(4) : 131-135, 2013 Note Food and feeding habits of Macrobrachium lar (Decapoda, Palaemonidae) from Andaman and Nicobar Islands, India S. N. SETHI1, NAGESH RAM2 AND V. VENKATESAN3 1Chennai Research Centre of Central Marine Fisheries Research Institute, 75 – Santhome High Road R. A. Puram, Chennai-600 028,Tamil Nadu, India 2Central Agricultural Research Institute (CARI), Port Blair-744 103, Andaman and Nicobar Islands, India 3Central Marine Fisheries Research Institute, Cochin -682 018, Kerala, India e-mail: [email protected] ABSTRACT The stomach content of the freshwater prawn Macrobrachium lar from the inland water bodies of Andaman and Nicobar Islands were analysed in relation to size, sex and season during the period from January to December 2008. Feeding habits of M. lar were studied using the index of preponderance and feeding intensity methods. Mainly organic detritus, supplemented by animal and plant materials formed the food in all stages of M. lar. The animal food items mainly consisted of aquatic insects, polychaetes, other crustaceans, fish, molluscs and zooplankton. The plant matter was chiefly composed of fragments of aquatic plants, planktonic algae and diatoms. Variation in diet in relation to size, sex or season were found insignificant. Though, all size groups of this species feed animal and plant materials, it exhibited slight preference for animal food especially in larger size groups. Females appear to feed more actively than the males. Feeding intensity was more in monsoon season compared to dry season. Results indicate that the adults are mostly predators of slow moving benthic invertebrates rather than detritus feeders/scavengers. -

WSR Vol 6 for 508 Pdf.Indd
Coastal and Estuarine Hazardous Waste Site Reports Editors J. Gardiner1, B. Azzato2, M. Jacobi1 1NOAA/OR&R/Coastal Protection and Restoration Division 2Azzato Communications Authors M. Hilgart, S. Duncan, S. Pollock Ridolfi Engineers Inc. NOAA National Oceanic and Atmospheric Administration NOS NOAA’s Ocean Service OR&R Office of Response and Restoration CPRD Coastal Protection and Restoration Division 7600 Sand Point Way NE Seattle, Washington 98115 September 30, 2004 Coastal and Estuarine Hazardous Waste Site Reports Reviewers K. Finkelstein1, M. Geddes2, M. Gielazyn1, R. Gouguet1, R. Mehran1 1NOAA/OR&R/Coastal Protection and Restoration Division 2Genwest Systems Graphics K. Galimanis 4 Point Design NOAA National Oceanic and Atmospheric Administration NOS NOAA’s Ocean Service OR&R Office of Response and Restoration CPRD Coastal Protection and Restoration Division 7600 Sand Point Way NE Seattle, Washington 98115 September 30, 2004 PLEASE CITE AS: J. Gardiner, B. Azzato and M. Jacobi, editors. 2004. Coastal and Estuarine Hazardous Waste Site Reports, September 30, 2004. Seattle: Coastal Protection and Restoration Division, Office of Response and Restoration, National Oceanic and Atmospheric Administration. 130 pp. v Contents Acronyms and abbreviations vii Introduction ix EPA Region 1 Callahan Mining Corp 1 Brooksville (Cape Rosier), Maine EPA Region 2 Diamond Head Oil Refinery Div. 11 Kearny, New Jersey MacKenzie Chemical Works 21 Central Islip, New York Pesticide Warehouse III 31 Manatí, Puerto Rico EPA Region 4 Davis Timber Company 41 Hattiesburg, -

Universidad De Costa Rica Facultad De Ciencias Escuela De Biologia
UNIVERSIDAD DE COSTA RICA FACULTAD DE CIENCIAS ESCUELA DE BIOLOGIA Optando por el grado académico de Licenciatura en Biología con énfasis en Recursos Acuáticos Morfometría y reproducción de tres especies langostinos de la vertiente del Pacífico de Costa Rica: Macrobrachium panamense, M. americanum y M. tenellum (Decapoda: Palaemonidae). Yurlandy Gutiérrez Jara Cédula 1-1057-0627 Carné: 985134 Miembros del comité Dr. Ingo Wehrtmann (Director de Tesis) M.Sc. Gerardo Umaña (Lector) M.Sc. Monika Springer (Lectora) MIEMBROS DEL COMITÉ REVISOR Firma: __________________________ Dr. Ingo Wehrtmann Director de Tesis Firma: __________________________ M.Sc. Gerardo Umaña Lector Firma: __________________________ M.Sc. Monika Springer Lectora Firma: __________________________ Dra. Virginia Solís Alvarado Presidenta del tribunal Firma: __________________________ Dr. Paul Hanson Revisor Externo Firma: __________________________ Biol. Yurlandy Gutiérrez Jara Postulante II Este trabajo esta dedicado con todo mi amor: a mi esposo Rólier Lara y mi hermoso hijo Matias Lara Gutiérrez A mi madre Virginia Jara Mis hermanas: Montserrath, Daniela y a mi sobrina Sophi Y con mucho cariño a mi hermana mayor Layin que desde el cielo siempre me cuida y guía TQM. III AGRADECIMIENTOS Agradezco a los profesores: Ingo por su ayuda y guía en el desarrollo de mi tesis. A Monika por sus valiosas sugerencias y a Don Gerardo por su colaboración, apoyo y formación en trabajo de campo. Además a todas las secres de Biología por su apoyo. A Jeffrey Sibaja por su guía en la utilización del programa estadístico, para la elaboración de pruebas. A la empresa Rainbow por su aporte económico en la logística del trabajo de campo, compra de equipo y viáticos utilizados.