Rainfall Correlates of Bird Population Fluctuations in a Puerto Rican Dry Forest: a Nine Year Study
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bird Checklist Guánica Biosphere Reserve Puerto Rico
United States Department of Agriculture BirD CheCklist Guánica Biosphere reserve Puerto rico Wayne J. Arendt, John Faaborg, Miguel Canals, and Jerry Bauer Forest Service Research & Development Southern Research Station Research Note SRS-23 The Authors: Wayne J. Arendt, International Institute of Tropical Forestry, U.S. Department of Agriculture Forest Service, Sabana Field Research Station, HC 2 Box 6205, Luquillo, PR 00773, USA; John Faaborg, Division of Biological Sciences, University of Missouri, Columbia, MO 65211-7400, USA; Miguel Canals, DRNA—Bosque de Guánica, P.O. Box 1185, Guánica, PR 00653-1185, USA; and Jerry Bauer, International Institute of Tropical Forestry, U.S. Department of Agriculture Forest Service, Río Piedras, PR 00926, USA. Cover Photos Large cover photograph by Jerry Bauer; small cover photographs by Mike Morel. Product Disclaimer The use of trade or firm names in this publication is for reader information and does not imply endorsement by the U.S. Department of Agriculture of any product or service. April 2015 Southern Research Station 200 W.T. Weaver Blvd. Asheville, NC 28804 www.srs.fs.usda.gov BirD CheCklist Guánica Biosphere reserve Puerto rico Wayne J. Arendt, John Faaborg, Miguel Canals, and Jerry Bauer ABSTRACt This research note compiles 43 years of research and monitoring data to produce the first comprehensive checklist of the dry forest avian community found within the Guánica Biosphere Reserve. We provide an overview of the reserve along with sighting locales, a list of 185 birds with their resident status and abundance, and a list of the available bird habitats. Photographs of habitats and some of the bird species are included. -

The Journal of Caribbean Ornithology
THE J OURNAL OF CARIBBEAN ORNITHOLOGY SOCIETY FOR THE C ONSERVATION AND S TUDY OF C ARIBBEAN B IRDS S OCIEDAD PARA LA C ONSERVACIÓN Y E STUDIO DE LAS A VES C ARIBEÑAS ASSOCIATION POUR LA C ONSERVATION ET L’ E TUDE DES O ISEAUX DE LA C ARAÏBE 2005 Vol. 18, No. 1 (ISSN 1527-7151) Formerly EL P ITIRRE CONTENTS RECUPERACIÓN DE A VES M IGRATORIAS N EÁRTICAS DEL O RDEN A NSERIFORMES EN C UBA . Pedro Blanco y Bárbara Sánchez ………………....................................................................................................................................................... 1 INVENTARIO DE LA A VIFAUNA DE T OPES DE C OLLANTES , S ANCTI S PÍRITUS , C UBA . Bárbara Sánchez ……..................... 7 NUEVO R EGISTRO Y C OMENTARIOS A DICIONALES S OBRE LA A VOCETA ( RECURVIROSTRA AMERICANA ) EN C UBA . Omar Labrada, Pedro Blanco, Elizabet S. Delgado, y Jarreton P. Rivero............................................................................... 13 AVES DE C AYO C ARENAS , C IÉNAGA DE B IRAMA , C UBA . Omar Labrada y Gabriel Cisneros ……………........................ 16 FORAGING B EHAVIOR OF T WO T YRANT F LYCATCHERS IN T RINIDAD : THE G REAT K ISKADEE ( PITANGUS SULPHURATUS ) AND T ROPICAL K INGBIRD ( TYRANNUS MELANCHOLICUS ). Nadira Mathura, Shawn O´Garro, Diane Thompson, Floyd E. Hayes, and Urmila S. Nandy........................................................................................................................................ 18 APPARENT N ESTING OF S OUTHERN L APWING ON A RUBA . Steven G. Mlodinow................................................................ -

<I>ANOLIS</I> LIZARDS in the FOOD WEBS of STRUCTURALLY
University of Tennessee, Knoxville TRACE: Tennessee Research and Creative Exchange Doctoral Dissertations Graduate School 12-2016 ASSESSING THE FUNCTIONAL SIMILARITY OF NATIVE AND INVASIVE ANOLIS LIZARDS IN THE FOOD WEBS OF STRUCTURALLY-SIMPLE HABITATS IN FLORIDA Nathan W. Turnbough University of Tennessee, Knoxville, [email protected] Follow this and additional works at: https://trace.tennessee.edu/utk_graddiss Part of the Terrestrial and Aquatic Ecology Commons Recommended Citation Turnbough, Nathan W., "ASSESSING THE FUNCTIONAL SIMILARITY OF NATIVE AND INVASIVE ANOLIS LIZARDS IN THE FOOD WEBS OF STRUCTURALLY-SIMPLE HABITATS IN FLORIDA. " PhD diss., University of Tennessee, 2016. https://trace.tennessee.edu/utk_graddiss/4174 This Dissertation is brought to you for free and open access by the Graduate School at TRACE: Tennessee Research and Creative Exchange. It has been accepted for inclusion in Doctoral Dissertations by an authorized administrator of TRACE: Tennessee Research and Creative Exchange. For more information, please contact [email protected]. To the Graduate Council: I am submitting herewith a dissertation written by Nathan W. Turnbough entitled "ASSESSING THE FUNCTIONAL SIMILARITY OF NATIVE AND INVASIVE ANOLIS LIZARDS IN THE FOOD WEBS OF STRUCTURALLY-SIMPLE HABITATS IN FLORIDA." I have examined the final electronic copy of this dissertation for form and content and recommend that it be accepted in partial fulfillment of the equirr ements for the degree of Doctor of Philosophy, with a major in Ecology and Evolutionary Biology. -

Eleutherodactylus Coqui Global Invasive Species Database (GISD)
FULL ACCOUNT FOR: Eleutherodactylus coqui Eleutherodactylus coqui System: Freshwater_terrestrial Kingdom Phylum Class Order Family Animalia Chordata Amphibia Anura Leptodactylidae Common name coqui (German), Puerto Rican treefrog (English, Puerto Rico), Caribbean tree frog (English), common coqui (English) Synonym Similar species Eleutherodactylus planirostris, Eleutherodactylus martinicensis Summary Eleutherodactylus coqui is a relatively small tree frog native to Puerto Rico. The frogs are quite adaptable to different ecological zones and elevations. Their loud call is the main reason they are considered a pest. E. coqui's mating call is its namesake, a high- pitched, two-note \"co-qui\" (ko-kee') which attains nearly 100 decibels at 0.5 metres. E. coqui have a voracious appetite and there is concern in Hawai‘i, where it has been introduced, that E. coqui may put Hawai‘i’s endemic insect and spider species at risk and compete with endemic birds and other native fauna which rely on insects for food. view this species on IUCN Red List Species Description Eleutherodactylus coqui is described as a relatively small treefrog. In Puerto Rico, mature calling males and \"parental males\" (males guarding a clutch) average about 34mm in length from snout to vent (snout-vent length, or SVL), while mature egg-laying females average about 41mm SVL. Like the true treefrogs (family Hylidae), E. coqui have well developed pads at the end of each toe that are used for sticking to surfaces. E. coqui are extremely variable in colouration. The dorsum (upper surface) is generally grey or grey-brown and uniform in colour, or may have either a dark \"M\" shape between the shoulders, two broad, light dorso-lateral bars (from the snout, through to the eye, to the axila of the rear legs) bordered with black spots and/or a light bar on top of the head between the eyes and a light underside stippled with brown (Campbell, 2000). -

Distribution, Ecology, and Life History of the Pearly-Eyed Thrasher (Margarops Fuscatus)
Adaptations of An Avian Supertramp: Distribution, Ecology, and Life History of the Pearly-Eyed Thrasher (Margarops fuscatus) Chapter 6: Survival and Dispersal The pearly-eyed thrasher has a wide geographical distribution, obtains regional and local abundance, and undergoes morphological plasticity on islands, especially at different elevations. It readily adapts to diverse habitats in noncompetitive situations. Its status as an avian supertramp becomes even more evident when one considers its proficiency in dispersing to and colonizing small, often sparsely The pearly-eye is a inhabited islands and disturbed habitats. long-lived species, Although rare in nature, an additional attribute of a supertramp would be a even for a tropical protracted lifetime once colonists become established. The pearly-eye possesses passerine. such an attribute. It is a long-lived species, even for a tropical passerine. This chapter treats adult thrasher survival, longevity, short- and long-range natal dispersal of the young, including the intrinsic and extrinsic characteristics of natal dispersers, and a comparison of the field techniques used in monitoring the spatiotemporal aspects of dispersal, e.g., observations, biotelemetry, and banding. Rounding out the chapter are some of the inherent and ecological factors influencing immature thrashers’ survival and dispersal, e.g., preferred habitat, diet, season, ectoparasites, and the effects of two major hurricanes, which resulted in food shortages following both disturbances. Annual Survival Rates (Rain-Forest Population) In the early 1990s, the tenet that tropical birds survive much longer than their north temperate counterparts, many of which are migratory, came into question (Karr et al. 1990). Whether or not the dogma can survive, however, awaits further empirical evidence from additional studies. -

Puerto Rico Comprehensive Wildlife Conservation Strategy 2005
Comprehensive Wildlife Conservation Strategy Puerto Rico PUERTO RICO COMPREHENSIVE WILDLIFE CONSERVATION STRATEGY 2005 Miguel A. García José A. Cruz-Burgos Eduardo Ventosa-Febles Ricardo López-Ortiz ii Comprehensive Wildlife Conservation Strategy Puerto Rico ACKNOWLEDGMENTS Financial support for the completion of this initiative was provided to the Puerto Rico Department of Natural and Environmental Resources (DNER) by U.S. Fish and Wildlife Service (USFWS) Federal Assistance Office. Special thanks to Mr. Michael L. Piccirilli, Ms. Nicole Jiménez-Cooper, Ms. Emily Jo Williams, and Ms. Christine Willis from the USFWS, Region 4, for their support through the preparation of this document. Thanks to the colleagues that participated in the Comprehensive Wildlife Conservation Strategy (CWCS) Steering Committee: Mr. Ramón F. Martínez, Mr. José Berríos, Mrs. Aida Rosario, Mr. José Chabert, and Dr. Craig Lilyestrom for their collaboration in different aspects of this strategy. Other colleagues from DNER also contributed significantly to complete this document within the limited time schedule: Ms. María Camacho, Mr. Ramón L. Rivera, Ms. Griselle Rodríguez Ferrer, Mr. Alberto Puente, Mr. José Sustache, Ms. María M. Santiago, Mrs. María de Lourdes Olmeda, Mr. Gustavo Olivieri, Mrs. Vanessa Gautier, Ms. Hana Y. López-Torres, Mrs. Carmen Cardona, and Mr. Iván Llerandi-Román. Also, special thanks to Mr. Juan Luis Martínez from the University of Puerto Rico, for designing the cover of this document. A number of collaborators participated in earlier revisions of this CWCS: Mr. Fernando Nuñez-García, Mr. José Berríos, Dr. Craig Lilyestrom, Mr. Miguel Figuerola and Mr. Leopoldo Miranda. A special recognition goes to the authors and collaborators of the supporting documents, particularly, Regulation No. -

Guide to Theecological Systemsof Puerto Rico
United States Department of Agriculture Guide to the Forest Service Ecological Systems International Institute of Tropical Forestry of Puerto Rico General Technical Report IITF-GTR-35 June 2009 Gary L. Miller and Ariel E. Lugo The Forest Service of the U.S. Department of Agriculture is dedicated to the principle of multiple use management of the Nation’s forest resources for sustained yields of wood, water, forage, wildlife, and recreation. Through forestry research, cooperation with the States and private forest owners, and management of the National Forests and national grasslands, it strives—as directed by Congress—to provide increasingly greater service to a growing Nation. The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, age, disability, and where applicable sex, marital status, familial status, parental status, religion, sexual orientation genetic information, political beliefs, reprisal, or because all or part of an individual’s income is derived from any public assistance program. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at (202) 720-2600 (voice and TDD).To file a complaint of discrimination, write USDA, Director, Office of Civil Rights, 1400 Independence Avenue, S.W. Washington, DC 20250-9410 or call (800) 795-3272 (voice) or (202) 720-6382 (TDD). USDA is an equal opportunity provider and employer. Authors Gary L. Miller is a professor, University of North Carolina, Environmental Studies, One University Heights, Asheville, NC 28804-3299. -

Have Bird Distributions Shifted Along an Elevational Gradient on a Tropical Mountain?
Received: 15 May 2017 | Revised: 13 September 2017 | Accepted: 28 September 2017 DOI: 10.1002/ece3.3520 ORIGINAL RESEARCH Have bird distributions shifted along an elevational gradient on a tropical mountain? Marconi Campos-Cerqueira1 | Wayne J. Arendt2 | Joseph M. Wunderle Jr2 | T. Mitchell Aide1 1University of Puerto Rico-Río Piedras, San Juan, Puerto Rico Abstract 2International Institute of Tropical An upward shift in elevation is one of the most conspicuous species responses to cli- Forestry, USDA Forest Service, Sabana Field mate change. Nevertheless, downward shifts and, apparently, the absences of Research Station, Luquillo, Puerto Rico response have also been recently reported. Given the growing evidence of multiple Correspondence responses of species distributions due to climate change and the paucity of studies in Marconi Campos-Cerqueira, University of Puerto Rico-Río Piedras, San Juan, Puerto the tropics, we evaluated the response of a montane bird community to climate Rico. change, without the confounding effects of land- use change. To test for elevational Email: [email protected] shifts, we compared the distribution of 21 avian species in 1998 and 2015 using Funding Information MCC was supported by the fellowship occupancy models. The historical data set was based on point counts, whereas the “Science Without Borders” from the contemporary data set was based on acoustic monitoring. We detected a similar num- Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) from Brazil ber of species in historical (36) and contemporary data sets (33). We show an overall (8933/13-8). pattern of no significant change in range limits for most species, although there was a significant shift in the range limit of eight species (38%). -

Dear Eastbay Volunteer
Area Search Species Codes Species Codes Please use the species codes on the list below. The full list is online at: http://www.pwrc.usgs.gov/bbl/manual/bandsize.htm Species Numbers, Common Names, Alpha Codes SPECIES COMMON NAME ALPHA CODE 001.0 Western Grebe WEGR 001.1 Clark's Grebe CLGR 002.0 Red-necked Grebe RNGR 003.0 Horned Grebe HOGR 004.0 Eared Grebe EAGR 005.0 Least Grebe LEGR 006.0 Pied-billed Grebe PBGR 007.0 Common Loon COLO 008.0 Yellow-billed Loon YBLO 009.0 Arctic Loon ARLO 010.0 Pacific Loon PALO 011.0 Red-throated Loon RTLO 012.0 Tufted Puffin TUPU 013.0 Atlantic Puffin ATPU 014.0 Horned Puffin HOPU 015.0 Rhinoceros Auklet RHAU 016.0 Cassin's Auklet CAAU 017.0 Parakeet Auklet PAAU 018.0 Crested Auklet CRAU 019.0 Whiskered Auklet WHAU 020.0 Least Auklet LEAU 021.0 Ancient Murrelet ANMU 023.0 Marbled Murrelet MAMU 023.1 Long-billed Murrelet LBMU 024.0 Kittlitz's Murrelet KIMU 025.0 Xantus's Murrelet XAMU 026.0 Craveri's Murrelet CRMU 027.0 Black Guillemot BLGU 029.0 Pigeon Guillemot PIGU 030.0 Common Murre COMU 031.0 Thick-billed Murre TBMU 032.0 Razorbill RAZO 034.0 Dovekie DOVE 035.0 Great Skua GRSK 035.2 South Polar Skua SPSK 036.0 Pomarine Jaeger POJA 037.0 Parasitic Jaeger PAJA 038.0 Long-tailed Jaeger LTJA 039.0 Ivory Gull IVGU 039.1 Magellan Gull MAGU 040.0 Black-legged Kittiwake BLKI 041.0 Red-legged Kittiwake RLKI 042.0 Glaucous Gull GLGU 043.0 Iceland Gull ICGU 043.1 Thayer's Gull THGU 044.0 Glaucous-winged Gull GWGU 044.6 Hybrid Gull HYGU 047.0 Great Black-backed Gull GBBG 048.0 Slaty-backed Gull SBGU 049.0 Western -

The Effects of Hurricanes on Birds, with Special Reference to Caribbean Islands
Bird Conservation International (1993) 3:319-349 The effects of hurricanes on birds, with special reference to Caribbean islands JAMES W. WILEY and JOSEPH M. WUNDERLE, JR. Summary Cyclonic storms, variously called typhoons, cyclones, or hurricanes (henceforth, hurricanes), are common in many parts of the world, where their frequent occurrence can have both direct and indirect effects on bird populations. Direct effects of hurricanes include mortality from exposure to hurricane winds, rains, and storm surges, and geo- graphic displacement of individuals by storm winds. Indirect effects become apparent in the storm's aftermath and include loss of food supplies or foraging substrates; loss of nests and nest or roost sites; increased vulnerability to predation; microclimate changes; and increased conflict with humans. The short-term response of bird populations to hurricane damage, before changes in plant succession, includes shifts in diet, foraging sites or habitats, and reproductive changes. Bird populations may show long-term responses to changes in plant succession as second-growth vegetation increases in storm- damaged old-growth forests. The greatest stress of a hurricane to most upland terrestrial bird populations occurs after its passage rather than during its impact. The most important effect of a hurricane is the destruction of vegetation, which secondarily affects wildlife in the storm's after- math. The most vulnerable terrestrial wildlife populations have a diet of nectar, fruit, or seeds; nest, roost, or forage on large old trees; require a closed forest canopy; have special microclimate requirements and/or live in a habitat in which vegetation has a slow recovery rate. Small populations with these traits are at greatest risk to hurricane-induced extinction, particularly if they exist in small isolated habitat fragments. -
![Inferring Habitat Quality and Habitat Selection Using Static Site Occupancy Models. Arxiv:1607.05175V1 [Q-Bio.PE] 18 Jul 2016](https://docslib.b-cdn.net/cover/6803/inferring-habitat-quality-and-habitat-selection-using-static-site-occupancy-models-arxiv-1607-05175v1-q-bio-pe-18-jul-2016-2606803.webp)
Inferring Habitat Quality and Habitat Selection Using Static Site Occupancy Models. Arxiv:1607.05175V1 [Q-Bio.PE] 18 Jul 2016
Inferring habitat quality and habitat selection using static site occupancy models. Philip T. Patton∗a, Krishna Pacificia and Jaime Collazoa,b aProgram in Fisheries, Wildlife, and Conservation Biology, Department of Applied Ecology, North Carolina State University, USA bU.S. Geological Survey, North Carolina Cooperative Fish and Wildlife Research Unit, North Carolina State University, USA Abstract When evaluating the ecological value of land use within a landscape, investigators typically rely on measures of habitat selection and habitat quality. Traditional measures of habitat selection and habitat quality require data from resource inten- sive study designs (e.g., telemetry, mark{recapture, or multi{season point counts). Often, managers must evaluate ecological value despite only having data from less resource intensive study designs. In this paper, we use occupancy data to mea- sure habitat quality and habitat selection response for the Puerto Rican Vireo, an endemic songbird whose population growth is depressed by brood parasitism from the Shiny Cowbird. We were interested in how vireo habitat quality and vireo habitat selection response varied among three land uses|forest, shaded cof- fee plantations, and sun coffee—in a coffee growing region in Puerto Rico. We estimated vireo occupancy probability as a measure of habitat selection, and the probability of cowbird occurrence given vireo presence as a measure of habitat quality. To estimate the latter, we explored different ways of incorporating host information into joint occurrence models. These included the conditional occur- rence model (Waddle et al., 2010), a two-stage hierarchical Bayesian model that propagates the joint uncertainty from the vireo and the cowbird through to es- timates of co-occurrence, and a commonly used model that includes the na¨ıve occurrence of the host as a covariate. -
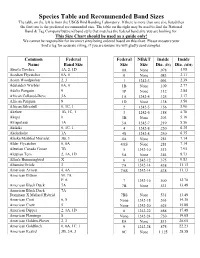
Species Table and Recommended Band Sizes the Table on the Left Is from the USGS Bird Banding Laboratory
Species Table and Recommended Band Sizes The table on the left is from the USGS Bird Banding Laboratory. If there is more than one size listed then the first one is the preferred recommended size. The table on the right may be used to find the National Band & Tag Company butt-end band style that matches the federal band size you are looking for. This Size Chart should be used as a guide only! We cannot be responsible for incorrect sizes being ordered based on this chart. Please measure your bird’s leg for accurate sizing, if you are unsure we will gladly send samples. Common Federal Federal NB&T Inside Inside Name Band Size Size Size Dia. (IN) Dia. (MM) Abert's Towhee 1A, 2, 1D 0A None .078 1.98 Acadian Flycatcher 0A, 0 0 None .083 2.11 Acorn Woodpecker 2, 3 1 1242-3 .094 2.39 Adelaide's Warbler 0A, 0 1B None .109 2.77 Adelie Penguin 9 1P None .112 2.84 African Collared-Dove 3A 1A 1242-4 .125 3.17 African Penguin 9 1D None .138 3.50 African Silverbill 0, 1C, 1 2 1242-5 .156 3.96 Akekee 1B, 1C, 1 3 1242-6 .188 4.78 Akepa 0 3B None .203 5.16 Akiapolaau 1A 3A 1242-7 .219 5.56 Akikiki 0, 1C, 1 4 1242-8 .250 6.35 Akohekohe 1A 4S 1242-8 .250 6.35 Alaska Marbled Murrelet 3B, 3 4A None .281 7.14 Alder Flycatcher 0, 0A 4AS None .281 7.14 Aleutian Canada Goose 7B 5 1242-10 .313 7.95 Aleutian Tern 2, 1A, 1D 5A None .344 8.73 Allen's Hummingbird X 6 1242-12 .375 9.53 Altamira Oriole 3 7A 1242-14 .438 11.13 American Avocet 4, 4A 7AS 1242-14 .438 11.13 American Bittern M: 7A F: 6 7 1242-16 .500 12.70 American Black Duck 7A 7B None .531 13.49 American