Inferring Habitat Quality and Habitat Selection Using Static Site Occupancy Models. Arxiv:1607.05175V1 [Q-Bio.PE] 18 Jul 2016
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Bird Checklist Guánica Biosphere Reserve Puerto Rico
United States Department of Agriculture BirD CheCklist Guánica Biosphere reserve Puerto rico Wayne J. Arendt, John Faaborg, Miguel Canals, and Jerry Bauer Forest Service Research & Development Southern Research Station Research Note SRS-23 The Authors: Wayne J. Arendt, International Institute of Tropical Forestry, U.S. Department of Agriculture Forest Service, Sabana Field Research Station, HC 2 Box 6205, Luquillo, PR 00773, USA; John Faaborg, Division of Biological Sciences, University of Missouri, Columbia, MO 65211-7400, USA; Miguel Canals, DRNA—Bosque de Guánica, P.O. Box 1185, Guánica, PR 00653-1185, USA; and Jerry Bauer, International Institute of Tropical Forestry, U.S. Department of Agriculture Forest Service, Río Piedras, PR 00926, USA. Cover Photos Large cover photograph by Jerry Bauer; small cover photographs by Mike Morel. Product Disclaimer The use of trade or firm names in this publication is for reader information and does not imply endorsement by the U.S. Department of Agriculture of any product or service. April 2015 Southern Research Station 200 W.T. Weaver Blvd. Asheville, NC 28804 www.srs.fs.usda.gov BirD CheCklist Guánica Biosphere reserve Puerto rico Wayne J. Arendt, John Faaborg, Miguel Canals, and Jerry Bauer ABSTRACt This research note compiles 43 years of research and monitoring data to produce the first comprehensive checklist of the dry forest avian community found within the Guánica Biosphere Reserve. We provide an overview of the reserve along with sighting locales, a list of 185 birds with their resident status and abundance, and a list of the available bird habitats. Photographs of habitats and some of the bird species are included. -

The Journal of Caribbean Ornithology
THE J OURNAL OF CARIBBEAN ORNITHOLOGY SOCIETY FOR THE C ONSERVATION AND S TUDY OF C ARIBBEAN B IRDS S OCIEDAD PARA LA C ONSERVACIÓN Y E STUDIO DE LAS A VES C ARIBEÑAS ASSOCIATION POUR LA C ONSERVATION ET L’ E TUDE DES O ISEAUX DE LA C ARAÏBE 2005 Vol. 18, No. 1 (ISSN 1527-7151) Formerly EL P ITIRRE CONTENTS RECUPERACIÓN DE A VES M IGRATORIAS N EÁRTICAS DEL O RDEN A NSERIFORMES EN C UBA . Pedro Blanco y Bárbara Sánchez ………………....................................................................................................................................................... 1 INVENTARIO DE LA A VIFAUNA DE T OPES DE C OLLANTES , S ANCTI S PÍRITUS , C UBA . Bárbara Sánchez ……..................... 7 NUEVO R EGISTRO Y C OMENTARIOS A DICIONALES S OBRE LA A VOCETA ( RECURVIROSTRA AMERICANA ) EN C UBA . Omar Labrada, Pedro Blanco, Elizabet S. Delgado, y Jarreton P. Rivero............................................................................... 13 AVES DE C AYO C ARENAS , C IÉNAGA DE B IRAMA , C UBA . Omar Labrada y Gabriel Cisneros ……………........................ 16 FORAGING B EHAVIOR OF T WO T YRANT F LYCATCHERS IN T RINIDAD : THE G REAT K ISKADEE ( PITANGUS SULPHURATUS ) AND T ROPICAL K INGBIRD ( TYRANNUS MELANCHOLICUS ). Nadira Mathura, Shawn O´Garro, Diane Thompson, Floyd E. Hayes, and Urmila S. Nandy........................................................................................................................................ 18 APPARENT N ESTING OF S OUTHERN L APWING ON A RUBA . Steven G. Mlodinow................................................................ -

<I>ANOLIS</I> LIZARDS in the FOOD WEBS of STRUCTURALLY
University of Tennessee, Knoxville TRACE: Tennessee Research and Creative Exchange Doctoral Dissertations Graduate School 12-2016 ASSESSING THE FUNCTIONAL SIMILARITY OF NATIVE AND INVASIVE ANOLIS LIZARDS IN THE FOOD WEBS OF STRUCTURALLY-SIMPLE HABITATS IN FLORIDA Nathan W. Turnbough University of Tennessee, Knoxville, [email protected] Follow this and additional works at: https://trace.tennessee.edu/utk_graddiss Part of the Terrestrial and Aquatic Ecology Commons Recommended Citation Turnbough, Nathan W., "ASSESSING THE FUNCTIONAL SIMILARITY OF NATIVE AND INVASIVE ANOLIS LIZARDS IN THE FOOD WEBS OF STRUCTURALLY-SIMPLE HABITATS IN FLORIDA. " PhD diss., University of Tennessee, 2016. https://trace.tennessee.edu/utk_graddiss/4174 This Dissertation is brought to you for free and open access by the Graduate School at TRACE: Tennessee Research and Creative Exchange. It has been accepted for inclusion in Doctoral Dissertations by an authorized administrator of TRACE: Tennessee Research and Creative Exchange. For more information, please contact [email protected]. To the Graduate Council: I am submitting herewith a dissertation written by Nathan W. Turnbough entitled "ASSESSING THE FUNCTIONAL SIMILARITY OF NATIVE AND INVASIVE ANOLIS LIZARDS IN THE FOOD WEBS OF STRUCTURALLY-SIMPLE HABITATS IN FLORIDA." I have examined the final electronic copy of this dissertation for form and content and recommend that it be accepted in partial fulfillment of the equirr ements for the degree of Doctor of Philosophy, with a major in Ecology and Evolutionary Biology. -

Rainfall Correlates of Bird Population Fluctuations in a Puerto Rican Dry Forest: a Nine Year Study
WilsonBull., 96(4), 1984,pp. 575-593 RAINFALL CORRELATES OF BIRD POPULATION FLUCTUATIONS IN A PUERTO RICAN DRY FOREST: A NINE YEAR STUDY JOHN FAABORG, WAYNE J. ARENDT, AND MARK S. KAISER Long-term studies on the population dynamics of Neotropical bird communities have been primarily limited to Panama (see Karr et al. [ 19821 for a mainland site and Willis [1974] for Barro Colorado Island). An earlier paper (Faaborg 1982a) contained the first long-term population measurements from a West Indian island, specifically a seasonally-dry forest site in southwest Puerto Rico. This Syear study apparently spanned a population peak followed by drought conditions and a severe population decline. The effects of drought on total populations, membership in dif- ferent foraging guilds, and winter resident densities were discussed. We have continued these studies and here report on 9 years of banding and population monitoring activities in a single location. This allows us to expand our previous observations on relationships between rainfall patterns and population traits of guilds and species and expose the data to statistical analyses. We also document the attempted invasion of a new species (Elaenia martinica) into the Guanica Forest bird community. The possible meaning of these observations in terms of island equilibrium theory (MacArthur and Wilson 1967), long-term climatic patterns (Pregill and Olson 198 l), and community structure studies (Faaborg 1982b) is discussed. STUDY AREA AND METHODS This study was done in seasonallydry scrubin the Guanica Forest of southwesternPuerto Rico (see Terborgh and Faaborg [1973] for detailed habitat description with photographs). This habitat occurs on a coraline limestone and contains sclerophyllousforest typical of such sites throughout the West Indies (Beard 1949). -

Puerto Rico Comprehensive Wildlife Conservation Strategy 2005
Comprehensive Wildlife Conservation Strategy Puerto Rico PUERTO RICO COMPREHENSIVE WILDLIFE CONSERVATION STRATEGY 2005 Miguel A. García José A. Cruz-Burgos Eduardo Ventosa-Febles Ricardo López-Ortiz ii Comprehensive Wildlife Conservation Strategy Puerto Rico ACKNOWLEDGMENTS Financial support for the completion of this initiative was provided to the Puerto Rico Department of Natural and Environmental Resources (DNER) by U.S. Fish and Wildlife Service (USFWS) Federal Assistance Office. Special thanks to Mr. Michael L. Piccirilli, Ms. Nicole Jiménez-Cooper, Ms. Emily Jo Williams, and Ms. Christine Willis from the USFWS, Region 4, for their support through the preparation of this document. Thanks to the colleagues that participated in the Comprehensive Wildlife Conservation Strategy (CWCS) Steering Committee: Mr. Ramón F. Martínez, Mr. José Berríos, Mrs. Aida Rosario, Mr. José Chabert, and Dr. Craig Lilyestrom for their collaboration in different aspects of this strategy. Other colleagues from DNER also contributed significantly to complete this document within the limited time schedule: Ms. María Camacho, Mr. Ramón L. Rivera, Ms. Griselle Rodríguez Ferrer, Mr. Alberto Puente, Mr. José Sustache, Ms. María M. Santiago, Mrs. María de Lourdes Olmeda, Mr. Gustavo Olivieri, Mrs. Vanessa Gautier, Ms. Hana Y. López-Torres, Mrs. Carmen Cardona, and Mr. Iván Llerandi-Román. Also, special thanks to Mr. Juan Luis Martínez from the University of Puerto Rico, for designing the cover of this document. A number of collaborators participated in earlier revisions of this CWCS: Mr. Fernando Nuñez-García, Mr. José Berríos, Dr. Craig Lilyestrom, Mr. Miguel Figuerola and Mr. Leopoldo Miranda. A special recognition goes to the authors and collaborators of the supporting documents, particularly, Regulation No. -

Guide to Theecological Systemsof Puerto Rico
United States Department of Agriculture Guide to the Forest Service Ecological Systems International Institute of Tropical Forestry of Puerto Rico General Technical Report IITF-GTR-35 June 2009 Gary L. Miller and Ariel E. Lugo The Forest Service of the U.S. Department of Agriculture is dedicated to the principle of multiple use management of the Nation’s forest resources for sustained yields of wood, water, forage, wildlife, and recreation. Through forestry research, cooperation with the States and private forest owners, and management of the National Forests and national grasslands, it strives—as directed by Congress—to provide increasingly greater service to a growing Nation. The U.S. Department of Agriculture (USDA) prohibits discrimination in all its programs and activities on the basis of race, color, national origin, age, disability, and where applicable sex, marital status, familial status, parental status, religion, sexual orientation genetic information, political beliefs, reprisal, or because all or part of an individual’s income is derived from any public assistance program. (Not all prohibited bases apply to all programs.) Persons with disabilities who require alternative means for communication of program information (Braille, large print, audiotape, etc.) should contact USDA’s TARGET Center at (202) 720-2600 (voice and TDD).To file a complaint of discrimination, write USDA, Director, Office of Civil Rights, 1400 Independence Avenue, S.W. Washington, DC 20250-9410 or call (800) 795-3272 (voice) or (202) 720-6382 (TDD). USDA is an equal opportunity provider and employer. Authors Gary L. Miller is a professor, University of North Carolina, Environmental Studies, One University Heights, Asheville, NC 28804-3299. -

Dear Eastbay Volunteer
Area Search Species Codes Species Codes Please use the species codes on the list below. The full list is online at: http://www.pwrc.usgs.gov/bbl/manual/bandsize.htm Species Numbers, Common Names, Alpha Codes SPECIES COMMON NAME ALPHA CODE 001.0 Western Grebe WEGR 001.1 Clark's Grebe CLGR 002.0 Red-necked Grebe RNGR 003.0 Horned Grebe HOGR 004.0 Eared Grebe EAGR 005.0 Least Grebe LEGR 006.0 Pied-billed Grebe PBGR 007.0 Common Loon COLO 008.0 Yellow-billed Loon YBLO 009.0 Arctic Loon ARLO 010.0 Pacific Loon PALO 011.0 Red-throated Loon RTLO 012.0 Tufted Puffin TUPU 013.0 Atlantic Puffin ATPU 014.0 Horned Puffin HOPU 015.0 Rhinoceros Auklet RHAU 016.0 Cassin's Auklet CAAU 017.0 Parakeet Auklet PAAU 018.0 Crested Auklet CRAU 019.0 Whiskered Auklet WHAU 020.0 Least Auklet LEAU 021.0 Ancient Murrelet ANMU 023.0 Marbled Murrelet MAMU 023.1 Long-billed Murrelet LBMU 024.0 Kittlitz's Murrelet KIMU 025.0 Xantus's Murrelet XAMU 026.0 Craveri's Murrelet CRMU 027.0 Black Guillemot BLGU 029.0 Pigeon Guillemot PIGU 030.0 Common Murre COMU 031.0 Thick-billed Murre TBMU 032.0 Razorbill RAZO 034.0 Dovekie DOVE 035.0 Great Skua GRSK 035.2 South Polar Skua SPSK 036.0 Pomarine Jaeger POJA 037.0 Parasitic Jaeger PAJA 038.0 Long-tailed Jaeger LTJA 039.0 Ivory Gull IVGU 039.1 Magellan Gull MAGU 040.0 Black-legged Kittiwake BLKI 041.0 Red-legged Kittiwake RLKI 042.0 Glaucous Gull GLGU 043.0 Iceland Gull ICGU 043.1 Thayer's Gull THGU 044.0 Glaucous-winged Gull GWGU 044.6 Hybrid Gull HYGU 047.0 Great Black-backed Gull GBBG 048.0 Slaty-backed Gull SBGU 049.0 Western -
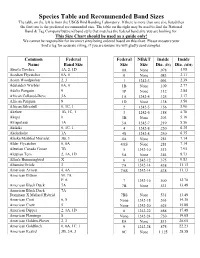
Species Table and Recommended Band Sizes the Table on the Left Is from the USGS Bird Banding Laboratory
Species Table and Recommended Band Sizes The table on the left is from the USGS Bird Banding Laboratory. If there is more than one size listed then the first one is the preferred recommended size. The table on the right may be used to find the National Band & Tag Company butt-end band style that matches the federal band size you are looking for. This Size Chart should be used as a guide only! We cannot be responsible for incorrect sizes being ordered based on this chart. Please measure your bird’s leg for accurate sizing, if you are unsure we will gladly send samples. Common Federal Federal NB&T Inside Inside Name Band Size Size Size Dia. (IN) Dia. (MM) Abert's Towhee 1A, 2, 1D 0A None .078 1.98 Acadian Flycatcher 0A, 0 0 None .083 2.11 Acorn Woodpecker 2, 3 1 1242-3 .094 2.39 Adelaide's Warbler 0A, 0 1B None .109 2.77 Adelie Penguin 9 1P None .112 2.84 African Collared-Dove 3A 1A 1242-4 .125 3.17 African Penguin 9 1D None .138 3.50 African Silverbill 0, 1C, 1 2 1242-5 .156 3.96 Akekee 1B, 1C, 1 3 1242-6 .188 4.78 Akepa 0 3B None .203 5.16 Akiapolaau 1A 3A 1242-7 .219 5.56 Akikiki 0, 1C, 1 4 1242-8 .250 6.35 Akohekohe 1A 4S 1242-8 .250 6.35 Alaska Marbled Murrelet 3B, 3 4A None .281 7.14 Alder Flycatcher 0, 0A 4AS None .281 7.14 Aleutian Canada Goose 7B 5 1242-10 .313 7.95 Aleutian Tern 2, 1A, 1D 5A None .344 8.73 Allen's Hummingbird X 6 1242-12 .375 9.53 Altamira Oriole 3 7A 1242-14 .438 11.13 American Avocet 4, 4A 7AS 1242-14 .438 11.13 American Bittern M: 7A F: 6 7 1242-16 .500 12.70 American Black Duck 7A 7B None .531 13.49 American -

List of Migratory Birds. Listed Alphabetically by Common (English) Group Names, with the Sci- (A) Legal Authority for This List
U.S. Fish and Wildlife Serv., Interior § 10.13 District of Columbia, Commonwealth sale, purchase, barter, exportation, and of Puerto Rico, American Samoa, U.S. importation of migratory birds. Virgin Islands, Guam, Commonwealth (c) What species are protected as migra- of the Northern Mariana Islands, Baker tory birds? Species protected as migra- Island, Howland Island, Jarvis Island, tory birds are listed in two formats to Johnston Atoll, Kingman Reef, Midway suit the varying needs of the user: Al- Atoll, Navassa Island, Palmyra Atoll, phabetically in paragraph (c)(1) of this and Wake Atoll, and any other terri- section and taxonomically in para- tory or possession under the jurisdic- graph (c)(2) of this section. Taxonomy tion of the United States. and nomenclature generally follow the Whoever means the same as person. 7th edition of the American Ornitholo- Wildlife means the same as fish or gists’ Union’s Check-list of North Amer- wildlife. ican birds (1998, as amended through 2007). For species not treated by the [38 FR 22015, Aug. 15, 1973, as amended at 42 AOU Check-list, we generally follow FR 32377, June 24, 1977; 42 FR 59358, Nov. 16, Monroe and Sibley’s A World Checklist 1977; 45 FR 56673, Aug. 25, 1980; 50 FR 52889, Dec. 26, 1985; 72 FR 48445, Aug. 23, 2007] of Birds (1993). (1) Alphabetical listing. Species are § 10.13 List of Migratory Birds. listed alphabetically by common (English) group names, with the sci- (a) Legal authority for this list. The entific name of each species following Migratory Bird Treaty Act (MBTA) in the common name. -

PUERTO RICO December 5-11, 2020
® field guides BIRDING TOURS WORLDWIDE [email protected] • 800•728•4953 ITINERARY PUERTO RICO December 5-11, 2020 The Puerto Rican Screech-Owl is one of the endemics we’ll see on the island. Unlike other screech-owls, this species does not have ear-tufts. Photograph by guide Tom Johnson. We include here information for those interested in the 2020 Field Guides Puerto Rico tour: ¾ a general introduction to the tour ¾ a description of the birding areas to be visited on the tour ¾ an abbreviated daily itinerary with some indication of the nature of each day’s outings Those who register for the tour will be sent this additional material: ¾ an annotated list of the birds recorded on a previous year’s Field Guides trip to the area, with comments by guide(s) on notable species or sightings (may be downloaded from our web site) ¾ a detailed information bulletin with important logistical information and answers to questions regarding accommodations, air arrangements, clothing, currency, customs and immigration, documents, health precautions, and personal items ¾ a reference list ¾ a Field Guides checklist for preparing for and keeping track of the birds we see on the tour ¾ after the conclusion of the tour, a list of birds seen on the tour Puerto Rico, easternmost and smallest of the Greater Antilles, is an exciting birding destination that harbors seventeen species of birds found nowhere else. Additionally, this US-associated Commonwealth features fine facilities and a good road system, making birding in Puerto Rico—from montane rainforests and elfin woods to mangrove swamps and dry coastal scrub—a comfortable and rewarding experience. -

Captive Wildlife Regulations, 2021, W-13.12 Reg 5
1 CAPTIVE WILDLIFE, 2021 W-13.12 REG 5 The Captive Wildlife Regulations, 2021 being Chapter W-13.12 Reg 5 (effective June 1, 2021). NOTE: This consolidation is not official. Amendments have been incorporated for convenience of reference and the original statutes and regulations should be consulted for all purposes of interpretation and application of the law. In order to preserve the integrity of the original statutes and regulations, errors that may have appeared are reproduced in this consolidation. 2 W-13.12 REG 5 CAPTIVE WILDLIFE, 2021 Table of Contents PART 1 PART 5 Preliminary Matters Zoo Licences and Travelling Zoo Licences 1 Title 38 Definition for Part 2 Definitions and interpretation 39 CAZA standards 3 Application 40 Requirements – zoo licence or travelling zoo licence PART 2 41 Breeding and release Designations, Prohibitions and Licences PART 6 4 Captive wildlife – designations Wildlife Rehabilitation Licences 5 Prohibition – holding unlisted species in captivity 42 Definitions for Part 6 Prohibition – holding restricted species in captivity 43 Standards for wildlife rehabilitation 7 Captive wildlife licences 44 No property acquired in wildlife held for 8 Licence not required rehabilitation 9 Application for captive wildlife licence 45 Requirements – wildlife rehabilitation licence 10 Renewal 46 Restrictions – wildlife not to be rehabilitated 11 Issuance or renewal of licence on terms and conditions 47 Wildlife rehabilitation practices 12 Licence or renewal term PART 7 Scientific Research Licences 13 Amendment, suspension, -

Puerto Rico 2017
Field Guides Tour Report Puerto Rico 2017 Mar 11, 2017 to Mar 17, 2017 Tom Johnson & Doug Gochfeld For our tour description, itinerary, past triplists, dates, fees, and more, please VISIT OUR TOUR PAGE. The breathtaking view of El Yunque National Forest from our lodge in the mountains of northeastern Puerto Rico. (Photo by guide Doug Gochfeld) “Wow” is the first word that jumps to mind when thinking of this year’s Field Guides tour to Puerto Rico. We had a splendid time circumnavigating the island and finding all 17 of the endemic bird species (and the other endemic taxa currently considered subspecies), with smashing views of most. We had six days jam-packed with goodness, including good birds, good food, and good humor. We started off very early on day 1 of the tour and drove from San Juan to Rio Abajo State Forest. We arrived before sunrise, and it was light enough for the dawn chorus to be starting but still dark enough that Puerto Rican Screech-Owls were still calling. Bill spotted one perched in a stand of bamboo where it was settled in to roost for the day. An early morning bonus here was Ruddy Quail-Dove. We then birded our way up the paved road until we arrived at a good vantage point from which to see one of the most endangered parrots in the world, the Puerto Rican Parrot. After a bit of a wait we connected with these loud, charismatic parrots, and there was much rejoicing. After Rio Abajo we went to Cambalache State Forest, where we were treated to excellent views of Puerto Rican Woodpecker, Puerto Rican Oriole, and Puerto Rican Flycatcher, as well as good views for some of Puerto Rican Lizard-Cuckoo.