The Northeast Greenland Shelf As a Potential Habitat for the Northeast Arctic Cod
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Værøy Kommune Elektro På Værøy Rådhuset, 8063 Værøy Tel
Værøy Municipality Text– Erling Skarv Johansen Photos– Leif Arne Olaussen and Erling Skarv Johansen A vibrant coastal community Life on Værøy In the middle of the open sea, near the fabled Moskstraumen Maelstrom, lies the island of Værøy – an active and intriguing fishing village with a rich history, midnight sun, white beaches, bird nesting cliffs, rapidly changing weather and a distinctive landscape. Surrounded by the sea, skerries and spectacular mountains, 750 people live here on the island, which is often referred to as „a hidden gem“. The landscape of Værøy could very well be prescribed as a cure for what ails you! A modern and vibrant island community Thriving cultural life Sørland is the island‘s main centre, and here you will find shops A wide range of clubs and associations ensures that there is and a café, local clothing designs and handicrafts, in addition to always something happening on Værøy at any given time. There a post office, school, day-care centre and doctor. Good transport is an activity for everyone here, regardless of age – whether it be connections are very important in a modern society, and we have spinning, skating, scouting, soccer, Christian Sports Contact, se- natural gas ferries and helicopters that go to Bodø every day. The nior dancing, fitness training, a youth club, a male choir, a female power grid on the island is buried underground together with fibre choir, the local radio or home arts and crafts! cable to the island, which provides very good coverage that opens up new opportunities. Stolt leverandør av tørr sk og lute sk fra Værøy www.brodreneberg.no www.brodreneberg.no People dare to take a chance here! On Værøy, there are many creative commercial actors, and the municipality is one of the country‘s most productive. -

Marine Science
ICES Journal of Marine Science ICES Journal of Marine Science (2014), 71(4), 907–908. doi:10.1093/icesjms/fst219 Contribution to the Themed Section: ‘Larval Fish Conference’ Introduction The early life history of fish—there is still a lot of work to do! Howard I. Browman* and Anne Berit Skiftesvik Downloaded from Institute of Marine Research, Austevoll Research Station, 5392 Storebø, Norway *Corresponding author: tel: +47 9886 0778; e-mail: [email protected] Browman, H. I., and Skiftesvik, A. B. 2014. The early life history of fish—there is still a lot of work to do! – ICES Journal of Marine Science, 71: 907–908. http://icesjms.oxfordjournals.org/ Received 6 November 2013; accepted 16 November 2013; advance access publication 9 January 2014. The themed set of articles that follows this introduction contains a selection of the papers that were presented at the 36th Annual Larval Fish Conference (ALFC), convened in Osøyro, Norway, 2–6 July 2012. The conference was organized around four theme sessions, three of which are represented with articles in this collection: “Assessing the relative contribution of different sources of mortality in the early life stages of fishes”; “The contribution of mechanistic, behavioural, and physiological studies on fish larvae to ecosystem models”; “Effects of oil and natural gas surveys, extraction activity and spills on fish early life stages”. Looking back at the main themes of earlier conferences about the early life history of fish reveals that they were not very different from those of ALFC2012. Clearly, we still have a lot of work to do on these and other topics related to the biology and ecology of fish early life stages. -

Maelstrom Kindle
MAELSTROM PDF, EPUB, EBOOK Peter Watts | 384 pages | 06 Jan 2009 | St Martins Press | 9780765320537 | English | New York, United States Maelstrom PDF Book Name: MAEL. Note that the 'protein existence' evidence does not give information on the accuracy or correctness of the sequence s displayed. GeneCards i. In the midst of a maelstrom , these technologies—among them social media, mobile apps, analytics, and cloud computing—help communities cope with the pandemic and learn crucial lessons. External Websites. Human Protein Atlas More PharmGKB i. After having been frozen in the waters they swam in, both reptiles remained in stasis until the ice ages began, thawing them out into a world ruled by mammals instead of reptiles. PeptideAtlas More Truthfully there were dozens upon dozens of reasons to have given up. Like his smaller companion Cretaceous, was incapable of speech and so communicated in loud growls, bellows, or roars. Account Orders My Account Sign out. Please enable JavaScript. I directed the 15th episode, which was right in the middle of a maelstrom of shooting and cutting The Divide. ExpressionAtlas i. You are using a version of browser that may not display all the features of this website. Bgee i. Learn More about maelstrom. Print Cite verified Cite. KEGG i. EvolutionaryTrace i. Participants 13 Sort alphabetically. RNAct i. The feeling of satisfaction was huge. TreeFam i. TreeFam database of animal gene trees More Examples of maelstrom in a Sentence She was caught in a maelstrom of emotions. Align Add to basket Added to basket This entry has 2 described isoforms and 1 potential isoform that is computationally mapped. -

Here Be Heathens: the Supernatural Image of Northern Fenno-Scandinavia in Pre-Modern Literature
Lokaverkefni til MA-gráðu í Norrænni trú Félagsvísindasvið Here be Heathens: The Supernatural Image of Northern Fenno-Scandinavia in Pre-Modern Literature Lyonel D. Perabo Norrænn trú Félags- og mannvísindadeild May 2016 Here be Heathens The Supernatural Image of Northern Fenno-Scandinavia in Pre-Modern Literature Lyonel D. Perabo Supervisor Terry Gunnell Faculty Representative Terry Gunnell Faculty of Social and Human Sciences School of Social Sciences University of Iceland Reykjavík, May 2016 Here be Heathens: The Supernatural Image of Northern Fenno-Scandinavia in Pre-Modern Literature Faculty of Social and Human Sciences School of Social Sciences University of Iceland Sæmundargata 2 101, Reykjavík Iceland Telephone: 525 4000 Bibliographic information: Lyonel D. Perabo, 2016, Here be Heathens: The Supernatural Image of Northern Fenno- Scandinavia in Pre-Modern Literature, Master’s thesis, Faculty of Social and Human Sciences, University of Iceland, pp. 222. Printing: Háskólaprent ehf. Tromsø, Norway: May 2016 "1 Abstract The present thesis involves a study of the various ways in which the Northernmost regions of Fenno-Scandinavia and their inhabitants were depicted as being associated with the supernatural in pre-Modern literature. Its findings are based on an exhaustive study of the numerous texts engaging with this subject, ranging from the Roman era to the publication of the Historia de Gentibus Septentrionalibus of Olaus Magnus in 1555. The thesis presents and analyses the most common supernatural motifs associated with this Far- Northern area, which include animal transformation, sorcery and pagan worship, as well as speculating about their origins, and analyzes the ways in which such ideas and images evolved, both over time and depending on the nature of the written sources in which they appear. -

Moskenes Guide Lofoten 2010
Side 1 MOSKENES GUIDE LOFOTEN 2010 MOSKENES GUIDE 2010 www.lofoten-info.no Velkommen til Moskenes Welcome to Moskenes Willkommen in Moskenes Side Kjære gjester og innbyggere, Vi har gleden av å ønske våre gjester velkommen til Moskenes. 2 Flakstad og Moskenes kan by på vakker natur, stupbratte fjell, hav og fiske, og et koselig og tradisjonsrikt miljø som vil danne rammen omkring ferien her hos oss. Vår reiselivsnæring er bygget på vår natur, kultur og egenart. Vi henstiller til innbyggere og feriegjester å hjelpe til med å ta vare på vår sårbare natur. Bevaring av lokalt særpreg ser vi som viktig for både fastboende og gjester. De fleste reiselivsbedriftene våre er små og eies av lokale familier som ønsker dere velkom- men til å bo og spise i originalt og tradisjonsrikt miljø. Vi ønsker våre gjester en riktig god ferie, og velkommen igjen. Dear visitor, It is my pleasure to wish you welcome to our community. Flakstad and Moskenes can offer magnificent scenery, soaring mountain peaks, the sea and fishing; a quaint environment rich in tradition - things which will form the framework of your holiday here in Lofoten. Our holiday resorts are based on our countryside, culture and way of life. We appeal to you, as a holiday guest, to help keep our vulnerable natural surroundings tidy and in tact. We consider the upkeep of our local traditions and identity as important to both locals and visitors alike. Most of our tourist businesses are small companies owned by families who welcome you to stay overnight and eat in an original and traditional environment Enjoy your holiday - and welcome back! Lieber Gast! Wir freuen uns, Sie in userem Gemeinden begrüßen zu dürfen. -

Models of Cross Shelf Transport Introduced by the Lofoten Maelstrom
Continental Shelf Research 22 (2002) 93–113 Models of cross shelf transport introduced by the Lofoten Maelstrom A. Ommundsen* Department of Mathematics, University of Oslo, P.O. Box 1053 Blindern, Oslo, Norway Received 1 July 1999; received in revised form 15 November 2000; accepted 27 April 2001 Abstract The drift of particles released in the tidal current (Moskstraumen) near Lofotodden, located at 671500N; 121500E; has been studied. The periodical tidal current combined with a background current is found to introduce a transport between Vestfjorden inside Lofotodden and the shelf outside. This particular transport may be important for the drift of cod eggs, larvae and other important biological and chemical tracers. Afine-resolution numerical model provided the Eulerian tidal current field based on which the particle trajectories are calculated using a Lagrangian particle tracking technique. The tracking algorithm was extended by a random-walk method to simulate small scale turbulence. An idealized current model of the area is also developed for use as a simplification tool to reveal the basic mechanisms of the drift and to demonstrate the sensibility of time stepping in numerical integration. r 2002 Elsevier Science Ltd. All rights reserved. Keywords: Maelstrom; Particle motion; Shelf dynamics; Tidal currents; Water exchange 1. Introduction outside Lofotodden is an example, running with strength up to 3–5 m=s: To make scientific well judged decisions regard- In an earlier work by Gjevik et al. (1997), a fine- ing the resources along the coast of Norway, better resolution numerical model for the tide around knowledge about how the tide influences the local Lofoten was developed. -

Modelling Wave-Current Interaction in the Lofoten Maelstrom
Modelling Wave-Current Interaction in the Lofoten Maelstrom Øyvind Saetra, Ana Carrasco and Trygve Halsne 22.12.2015 Norwegian Meteorological Institute Overview • The lofoten Maelstrom, locally named Moskstraumen • Intense tidal ocean current in Northern Norway • Classified as dangerous sailing area in the Pilot Guide for Norway • All physical information so far are based on eyewitness accounts - no existing instrumental observations available • Ships have been observed stationary at 10 knots (5 m/s) • Complicated sailing conditions due to surface currents often with opposing the local wave direction Here, we will present: • High-resolution ocean modelling (800m) • Observations from satellites (Optical and SAR) and photographs from land, collocated with numerical model results • Current and wave modelling - wave-current interaction 2 • Plans for deploying wave-current observing instrumets Moskstraumen - The Lofoten Maelstrom 3 Norwegian Meteorological Institute Moskstraumen - history and myths Carta Marina by Olaus Magnus, Venice 1539 • One of the worlds strongest tidal currents in open ocean areas (> 3m/s) • Described in Nordic tales as a magical millstone grinding salt on the bottom of the ocean • Showed as a gigantic eddy in Olaus Magnus’ map Carta Marina from 1539 • In international literature by Jules Verne Twenty Thousand Leagues Under the Sea, Edgar Allan Poe A descent into the Malström and Peter Dass, The trumpet of 4 Nordland Norwegian Meteorological Institute 5 Norwegian Meteorological Institute 6 Norwegian Meteorological Institute -
A High Resolution Tidal Model for the Area Around the Lofoten Islands,Northern Norway
Continental Shelf Research 22 (2002) 485–504 A high resolution tidal model for the area around The Lofoten Islands,northern Norway H. Moe,A. Ommundsen,B. Gjevik* Department of Mathematics, University of Oslo, Box 1053 Blindern, Oslo, Norway Received 07 April 2000; received in revised form 19 June 2001; accepted 28 June 2001 Abstract A depth-integrated numerical model with grid resolution 500 m has been used to simulate tides around the Lofoten Islands in northern Norway. The model spans more than 31 latitude and covers a sea area of approximately 1:2 Â 105 km2: The fine spatial resolution resolves the important fine scale features of the bottom topography on the shelf and the complex coastline with fjords and islands. Boundary conditions at the oceanic sides of the model domain are obtained by interpolation from a large-scale tidal model covering the Nordic Seas and implemented with the flow relaxation scheme (FRS). The semi-diurnal components M2; S2 and N2 and the diurnal component K1 are simulated. Harmonic constants for sea level are compared with observations from 21 stations. The best fit is found for the M2 component with a standard deviation between the observed and modelled amplitude and phase of 2:3 cm and 2:51; respectively. The standard deviation for the other smaller components ranges between 1.5–2:8 cm and 5.3–16:71: Current fields from the model are compared with observations in four locations: the Moskenes sound,the Gims y channel,the Tjeldsund channel and the Sortland channel. In the Sortland channel,the model predicts a dominant diurnal K1 current in agreement with observations. -

Biofokus-Rapport 2008-14
Ekstrakt Biofokus-rapport 2008-14 BioFokus har på oppdrag fra Fylkesmannen i Nordland, miljøvernavdelingen, Værøy Tittel kommune, Moskenes kom- mune og Direktoratet for na- Naturtypekartlegging i Moskenes og Værøy kommu- turforvaltning kvalitetssikret ner 2007 og nykartlagt naturtyper i Moskenes og Værøy kommu- ner. Det har totalt blitt re- Forfattere gistrert 38 lokaliteter hvorav Terje Blindheim, Jon Klepsland, Kjell Magne Olsen, 20 i Værøy og 18 i Moske- Tom H. Hofton og Kim Abel nes. Tidligere kartlegginger er gjennomgått og det er ut- ført nytt feltarbeid. Det er i Dato denne rapporten gitt en kort gjennomgang av status for 01.03.2009 naturtypekartleggingen i de to kommunene. Alle data er overlevert fylkesmannen for Antall sider bruk i naturbase. 31 sider Nøkkelord Publiseringstype Nordland Digitalt dokument (Pdf). Som digitalt dokument inneholder Værøy denne rapporten ”levende” linker. Moskenes Naturtypekartlegging Verdisetting Oppdragsgiver Flyvesandsområder Lofoten Fylkesmannen i Nordland, miljøvernavdelingen, Værøy kommune, Moskenes kommune og Direktoratet for natur- Omslag forvaltning FORSIDEBILDER (FOTO KIM ABEL) Øvre (Bakkesøte): Tilgjengelighet Midtre (Bunesfjorden): Dokumentet er offentlig tilgjengelig. Nedre (Sanddyner): Andre BioFokus rapporter kan lastes ned fra: http://biolitt.biofokus.no/rapporter/Litteratur.htm LAYOUT (OMSLAG) Blindheim Grafisk ISSN: 1504-6370 ISBN: 978-82-8209-043-8 BioFokus: Gaustadallèen 21, 0349 OSLO Telefon 2295 8598 E-post: [email protected] Web: www.biofokus.no Forord Stiftelsen Biofokus har på oppdrag fra Fylkesmannen i Nordland, miljøvernavdelingen, Vær- øy kommune, Moskenes kommune og Direktoratet for naturforvaltning foretatt en oppdate- ring av Moskenes og Værøy kommuner sine naturtypekart. Sveinung Råheim hos Fylkes- mannen har vært vår kontaktperson hos oppdragsgiver. Han skal ha takk for å ha vært til stor hjelp med effektivt å skaffe til veie digitalt og analogt kartgrunnlag, eksisterende in- formasjon, samt tips om transportmuligheter i kommunene. -
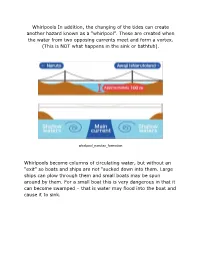
Whirlpools in Addition, the Changing of the Tides Can Create Another Hazard Known As a “Whirlpool”
Whirlpools In addition, the changing of the tides can create another hazard known as a “whirlpool”. These are created when the water from two opposing currents meet and form a vortex. (This is NOT what happens in the sink or bathtub). whirlpool_narutao_formation Whirlpools become columns of circulating water, but without an “exit” so boats and ships are not “sucked down into them. Large ships can plow through them and small boats may be spun around by them. For a small boat this is very dangerous in that it can become swamped – that is water may flood into the boat and cause it to sink. Whether or not the whirlpool appears and how rapidly it spins are a function of the tides. High tides and especially spring tides produce the most intense whirlpools. The figures given below are “highs”. In some cases, these have become tourist attractions with boats taking passengers out to see the whirlpool “up close” whirlpool_narutao_tourists The largest whirlpools are Saltstraumen (23 mph) in Norway, Whirlpool_Saltstraumen_Norway Moskstraumen (17.3) also in Norway, although there are those who think this is more an “eddy” than a whirl pool and Corryvreckan in Scotland found off the west coast between Jura and Scarpa. It has speeds of about 12 mph Whirlpool_Corryvreckan A dramatic encounter with Corryvreckan can be seen in the film I Know Where I’m Going. On the US/Canadian border lies Old Sow (17.1 mph) between Deer Island in New Brunswick and Moose Island, Eastport Maine. The US Coast Guard Station in Eastport regularly rescues boats that have gotten too close and do not have enough power to move them out of the current. -

Maine Tidal In-Stream Energy Conversion (TISEC): Survey and Characterization of Potential Project Sites
Maine Tidal In-Stream Energy Conversion (TISEC): Survey and Characterization of Potential Project Sites Project: EPRI North American Tidal Flow Power Feasibility Demonstration Project Phase: 1 – Project Definition Study Report: EPRI - TP- 003 ME Author: George Hagerman Co-Author: Roger Bedard Date: April 7, 2006 North America Tidal In Stream Energy Conversion Feasibility Study – Site Report - Maine ACKNOWLEDGEMENT The work described in this report was funded by the Maine Technology Initiative (MTI) who, in addition to providing funding for the EPRI Project Team, provided valuable in kind contributions to the project, especially in terms of James Atwell from the MTI Board of Directors Two special acknowledgements are also extended; one to Michael Mayhew of the Maine PUC and the other to Robert Judd, citizen of Lubec Maine and Sacramento, CA, for there tireless work in helping the EPRI team with local knowledge and resources. If, despite our best efforts, errors survive in the pages of this summary report or in our referenced technical reports, the fault rests entirely with the EPRI E2I Global Project Team. And if I inadvertently omitted the names of any persons or organizations that should have been acknowledged, I offer my sincere apologies. Roger Bedard DISCLAIMER OF WARRANTIES AND LIMITATION OF LIABILITIES This document was prepared by the organizations named below as an account of work sponsored or cosponsored by the Electric Power Research Institute Inc. (EPRI). Neither EPRI, any member of EPRI, any cosponsor, the organization (s) -

Uu.Diva-Portal.Org
http://uu.diva-portal.org This is an author produced version of a paper published in Renewable & Sustainable Energy Reviews. This paper has been peer-reviewed but does not include the final publisher proof-corrections or journal pagination. Citation for the published paper: Grabbe, Mårten; Lalander, Emilia; Lundin, Staffan; Leijon, Mats "A review of the tidal current energy resource in Norway" Renewable & Sustainable Energy Reviews, 2009, Vol. 13, Issue 8, pp. 1898-1909 URL: http://dx.doi.org/10.1016/j.rser.2009.01.026 Access to the published version may require subscription. A review of the tidal current energy resource in Norway Marten˚ Grabbe Emilia Lalander Staffan Lundin Mats Leijon The Swedish Centre for Renewable Electric Energy Conversion Division of Electricity, The Angstr˚ om¨ Laboratory Uppsala University P.O. Box 534, SE-751 21 Uppsala, Sweden Telephone: +46 (0)18 471 5843, Fax: +46 (0)18 471 5810 Email: [email protected] Website: http://www.el.angstrom.uu.se/ Abstract As interest in renewable energy sources is steadily on the rise, tidal current energy is receiving more and more attention from politicans, industrialists, and academics. In this article, the conditions for and potential of tidal currents as an energy resource in Norway are reviewed. There having been a relatively small amount of academic work published in this particular field, closely related topics such as the energy situation in Norway in general, the oceanography of the Norwegian coastline, and numerical models of tidal currents in Norwegian waters are also examined. Two published tidal energy resource assessments are reviewed and compared to a desktop study made specifically for this review based on available data in pilot books.