Lizards from Prehistoric Sites on Ebon Atoll, Marshall Islands Department
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Sustained Prehistoric Exploitation of a Marshall Islands Fishery
Sustained prehistoric exploitation of a Marshall Islands fishery: ichthyoarchaeological approaches, marine resource use, and human- environment interactions on Ebon Atoll Ariana Blaney Joan Lambrides BA (Hons) A thesis submitted for the degree of Doctor of Philosophy at The University of Queensland in 2017 School of Social Science Abstract Atolls are often characterised in terms of the environmental constraints and challenges these landscapes impose on sustained habitation, including: nutrient-poor soils and salt laden winds that impede plant growth, lack of perennial surface fresh water, limited terrestrial biodiversity, and vulnerability to extreme weather events and inundation since most atolls are only 2-3 m above sea level. Yet, on Ebon Atoll, Republic of the Marshall Islands in eastern Micronesia, the oceanside and lagoonside intertidal marine environments are expansive, with the reef area four times larger than the land area, supporting a diverse range of taxa. Given the importance of finfish resources in the Pacific, and specifically Ebon, this provided an ideal context for evaluating methods and methodological approaches for conducting Pacific ichthyoarchaeological analyses, and based on this assessment, implement high resolution and globally recognised approaches to investigate the spatial and temporal variation in the Ebon marine fishery. Variability in landscape use, alterations in the range of taxa captured, archaeological proxies of past climate stability, and the comparability of archaeological and ecological datasets were considered. Utilising a historical ecology approach, this thesis provides an analysis of the exploitation of the Ebon marine fishery from initial settlement to the historic period—two millennia of continuous occupation. The thesis demonstrated the importance of implementing high resolution methods and methodologies when considering long-term human interactions with marine fisheries. -

Statistical Yearbook, 2017
REPUBLIC OF THE MARSHALL ISLANDS STATISTICAL YEAR BOOK 2017 Economic Policy, Planning and Statistics Office (EPPSO) Office of the President Republic of the Marshall Islands RMI Statistical Yearbook, 2017 Statistical Yearbook 2017 Published by: Economic Policy, Planning and Statistics Office (EPPSO), Office of the President, Republic of the Marshall Islands Publication Year: June, 2018 Technical support was provided by Inclusive Growth Thematic cluster, UNDP, Pacific Office, Suva, Fiji Disclaimer The opinions expressed herein are those of the author and do not necessarily reflect the views of the UNDP or EPPSO. The pictures used in this publication are mostly taken from the Google search and some from the respective organization’s websites. EPPSO is not responsible if there is any violation of “copy right” issue related with any of them. 1 RMI Statistical Yearbook, 2017 TABLE OF CONTENTS PREFACE ...................................................................................................................................... 5 FOREWORD .................................................................................................................................. 6 LIST OF ACRONYMS ...................................................................................................................... 7 SUGGESTED NOTES PRIOR TO READING THIS PUBLICATION .......................................................... 10 BRIEF HISTORY OF REPUBLIC OF THE MARSHALL ISLANDS ............................................................. 12 REPUBLIC -

Atoll Research Bulletin No. 503 the Vascular Plants Of
ATOLL RESEARCH BULLETIN NO. 503 THE VASCULAR PLANTS OF MAJURO ATOLL, REPUBLIC OF THE MARSHALL ISLANDS BY NANCY VANDER VELDE ISSUED BY NATIONAL MUSEUM OF NATURAL HISTORY SMITHSONIAN INSTITUTION WASHINGTON, D.C., U.S.A. AUGUST 2003 Uliga Figure 1. Majuro Atoll THE VASCULAR PLANTS OF MAJURO ATOLL, REPUBLIC OF THE MARSHALL ISLANDS ABSTRACT Majuro Atoll has been a center of activity for the Marshall Islands since 1944 and is now the major population center and port of entry for the country. Previous to the accompanying study, no thorough documentation has been made of the vascular plants of Majuro Atoll. There were only reports that were either part of much larger discussions on the entire Micronesian region or the Marshall Islands as a whole, and were of a very limited scope. Previous reports by Fosberg, Sachet & Oliver (1979, 1982, 1987) presented only 115 vascular plants on Majuro Atoll. In this study, 563 vascular plants have been recorded on Majuro. INTRODUCTION The accompanying report presents a complete flora of Majuro Atoll, which has never been done before. It includes a listing of all species, notation as to origin (i.e. indigenous, aboriginal introduction, recent introduction), as well as the original range of each. The major synonyms are also listed. For almost all, English common names are presented. Marshallese names are given, where these were found, and spelled according to the current spelling system, aside from limitations in diacritic markings. A brief notation of location is given for many of the species. The entire list of 563 plants is provided to give the people a means of gaining a better understanding of the nature of the plants of Majuro Atoll. -

ATOLL RESEARCH BULLETIN No. Xi the Land Vegetation of Arno Atoll
ATOLL RESEARCH BULLETIN No. xi The Land Vegetation of Arno Atoll, Marshall Islands by William H. Hatheway Issued by THE PACIFIC SCIENCE EOARD National Academy of Sciences--National Research Council Washington, D.C. April 30, 1951 . he Land Vegetation of &no A$&,, ~arsh&l~slanda ' '' .. SCIENTIFIC DJVESTIGATIONS IN MICRONESIA Pacific Science Board National Research Council . Wi.llbm :I. Hatheway Haj:vw'd University Caibc:.c!&, ~lassachusetts~ . Jariuwy. 1953 ... , This report presents the results of botanical field uork done in the summer of 1.952 under the Coral ,!toll Project of the Pacis'ic Science Board of the Natj.onal Research Council. The work was supported by funds granted to the National Academy of Sclences by the Office of Naval Research. The fieldwork was carried out with the active assista.nce of the U. S. Navy Department, the Nilitary Air Transport Service, and the Trust Territory of the Pacific Islands. The miter wishes to express his thanks for help received from Harold Ja Coolidge, :Piis, Lenore Smi.th, and Miss Ernestine Akers. Mr. D. W, Gilfillan, District Ad:n*k&?ator of the Trust Terrctoiy at Xajuro, and Mro J, E. Tobin, staff ant:2sopolpgistj took especial in- terest in this stxdy and assisteci greatly. The help of Lt. Cmdr, CI IC, Brust, who arranged to have put a't the disposal of the author certain re- sources of the Ua S. Navy, is grateful!_y ackno~dledged, Special thanks are due the people of Arno, especially Kihej L., Felix J., Tobu, Lajibili., Jemeliic, Jibloic J., Abijai, Lakarim, jonag Nemot L*, Jajin F,, Bingel, Jakalu, Saburo, Jon, Lauken, Overton, and Likotalc* At Na.juro Raymond QeBrw., Konto S., and Bright 3eine helped me in many ways. -

Ground-Water Resources of the Laura Area, Majuro Atoll, Marshall Islands
GROUND-WATER RESOURCES OF THE LAURA AREA, MAJURO ATOLL, MARSHALL ISLANDS By Scott N. Hamlin and Stephen S. Anthony U.S. GEOLOGICAL SURVEY Water-Resources Investigations Report 87-4047 Prepared in cooperation with the REPUBLIC OF THE MARSHALL ISLANDS Honolulu, Hawaii 1987 UNITED STATES DEPARTMENT OF THE INTERIOR DONALD PAUL HODEL, Secretary GEOLOGICAL SURVEY Dallas L. Peck, Director For additional information Copies of this report write to: can be purchased from: District Chief, Hawaii District U.S. Geological Survey U.S. Geological Survey, WRD Books and Open-File Reports Section 300 Ala Moana Blvd., Rm. 6110 Federal Center, Bldg. 41 Honolulu, Hawai 96850 Box 25425 Denver, Colorado 80225 CONTENTS Page Abstract ------------------------------------------------------------ 1 Introduction -------------------------------------------------------- 2 Purpose and scope ---------------------------------------------- 5 Setting -------------------------------------------------------- 5 Previous investigations ---------------------------------------- 6 Acknowledgments ------------------------------------------------ 8 Methods of study ---------------------------------------------------- 8 Surface geophysical survey ------------------------------------- 8 Installation of driven-well network ---------------------------- 8 Test holes and collection of lithologic samples ---------------- 11 Collection of water samples ------------------------------------ 11 Measurements of water levels ----------------------------------- 13 Geohydrologic framework --------------------------------------------- -

Review and Evaluation of the Use of Tobacco, Alcohol and Opium in the Pre-Colonial Marshall Islands
THE JOHNSTONE CENTRE REPORT Nº 142 Review and Evaluation of the use of Tobacco, Alcohol and Opium in the pre-colonial Marshall Islands by Dirk H.R. Spennemann ALBURY 2000 © The author 2000. All rights reserved. The contents of this study are copyright in all countries subscribing to the Berne Convention. No parts of this book may be reproduced or transmitted in any form or by any means, electronic or mechanical, including photocopying, recording or by any information storage and retrieval system, without the written permission of the author, except where permitted by law. CIP Spennemann, Dirk H.R. 1958— Review and Evaluation of the use of Tobacco, Alcohol and Opium in the pre- colonial Marshall Islands- by Dirk H.R. Spennemann Albury, NSW: Charles Sturt University, The Johnstone Centre, 2000. 1 v., - (Report / Johnstone Centre of Parks, Recreation & Heritage, no. 142) I. Charles Sturt University. Johnstone Centre of Parks, Recreation & Heritage. II. Title. III. Series. Embargoed until 30 June 2001 Contents List of Figures....................................................................... vi List of Tables........................................................................ vi Introduction ......................................................................1 The Sources.......................................................................... 2 1 • Tobacco & Alcohol in the pre 19th Century Marshalls ................................................................23 Early Contact...................................................................... -

Destination: Marshall Islands. Video Guide. INSTITUTION Peace Corps, Washington, DC
DOCUMENT RESUME ED 354 198 SO 022 829 TITLE Destination: Marshall Islands. Video Guide. INSTITUTION Peace Corps, Washington, DC. Office of World Wise Schools. PUB DATE Jan 92 NOTE 82p.; For related documents, see SO 022 828-832. Accompanying videotape available from the producer; videotapes will be sent to the school librarian or media specialist. AVAILABLE FROM Peace Corps of the United States of America, Office of World Wise Schools, 1990 K Street, N.W., Washington, DC 20526. PUB TYPE Guides Classroom Use Teaching Guides (For Teacher)(052) EDRS PRICE MF01/PC04 Plus Postage. DESCRIPTORS Area Studies; Elementary Secondary Education; *Geographic Concepts; *Geography Instruction; *Multicultural Education; Social Studies; Thematic Approach; Videotape Recordings IDENTIFIERS Global Education; *Marshall Islands; *Peace Corps ABSTRACT This video guide was developed by the Peace Corps' Office of World Wise Schools. Activities that the guide describes are for use in a 3- to 5-day unit on one of the nations of Oceania, the Republic of the Marshall Islands. The activities are designed to provide students with opportunities to:(1) compr.re and contrast Marshallese and U.S. culture; and (2) relate the fundamental geographic themes of location, place, and movement with the history and culture of the Marshall Islands. Included in the guide are Level A, for grades three through five, Level B, for grades six through nine, and Level C, for grades 10 through 12. Each level includes worksheets and suggestions for using them. Fifty-seven references and a list of organizations that provided information are included. (LBG) *********************************************************************** Reproductions supplied by EDRS are the best that can be made from the original document. -

Atoll Research Bulletin No. 330 a Review of The
ATOLL RESEARCH BULLETIN NO. 330 A REVIEW OF THE NATURAL HISTORY OF THE MARSHALL ISLANDS BY F. RAYMOND FOSBERG ISSUED BY NATIONAL MUSEUM OF NATURAL HISTORY SMITHSONIAN INSTITUTION WASHINGTON, D.C., U.S.A. January 1990 A REVIEW OF THE NATURAL HISTORY OF THE MARSHALL ISLANDS F. RAYMOND FOSBERG This review was prepared on very short notice, to provide a summary of what is known to the reviewer at the time, June 1988, about the natural phenomena of the Marshall Islands. This was for the use of the members of the survay team sent to the Marshalls by the Environmentand Policy Institute of the East-West Center, Honolulu. Their mission was to investigate the remaining relatively natural areas and the extent of biodiversity in the new Republic of the Marshall Islands. The Marshall Archipelago has been the habitat of Aboriginal man for many hundreds, perhaps even thousands, of years. Hence there is very little, if any, undisturbed land remaining. Since the coming of the Europeans, in the 19th Century, and especially in the years 1940 to the present, the disturbance and change have been greatly intensified. On four of the Atolls, namely, Eniwetok, Bikini, Kwajalein, and Majuro, the alteration has been catastrophic. Change is so rapid that even a superficial account of what was observed during the early and mid 1950's seems worth placing on permanent record. This account is definitely not the results of a systematic literature search, but merely what is stores in the reviewer's head and in his notebooks, with such additions as are acknowkedged in the text. -

Pacific Islands
PUB. 126 SAILING DIRECTIONS (ENROUTE) ★ PACIFIC ISLANDS ★ Prepared and published by the NATIONAL GEOSPATIAL-INTELLIGENCE AGENCY Springfield, Virginia © COPYRIGHT 2014 BY THE UNITED STATES GOVERNMENT NO COPYRIGHT CLAIMED UNDER TITLE 17 U.S.C. 2014 ELEVENTH EDITION For sale by the Superintendent of Documents, U.S. Government Printing Office Internet: http://bookstore.gpo.gov Phone: toll free (866) 512-1800; DC area (202) 512-1800 Fax: (202) 512-2250 Mail Stop: SSOP, Washington, DC 20402-0001 III Preface 0.0 Pub. 126, Sailing Directions (Enroute) for Pacific Islands, 0.0 New editions of Sailing Directions are corrected through the Eleventh Edition, 2014, is issued for use in conjunction with date of the publication shown above. Important information to Pub. 120, Sailing Directions (Planning Guide) Pacific Ocean amend material in the publication is available as a Publication and Southeast Asia. Companion volumes are Pubs. 125 and Data Update (PDU) from the NGA Maritime Domain web site. 127. 0.0 Digital Nautical Charts 6, 12, and 13 provide electronic chart 0.0NGA Maritime Domain Website coverage for the area covered by this publication. http://msi.nga.mil/NGAPortal/MSI.portal 0.0 This publication has been corrected to 19 July 2014, including Notice to Mariners No. 29 of 2014. 0.0 0.0 Courses.—Courses are true, and are expressed in the same Explanatory Remarks manner as bearings. The directives “steer” and “make good” a course mean, without exception, to proceed from a point of 0.0 Sailing Directions are published by the National Geospatial- origin along a track having the identical meridianal angle as the Intelligence Agency (NGA), under the authority of Department designated course. -
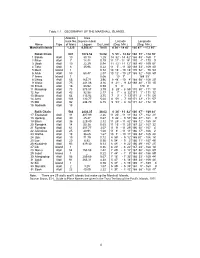
Table 1.1 GEOGRAPHY of the MARSHALL ISLANDS. Approxi- Area Mate No. (Square Miles) Latitude Longitude Name Type of Islets
Table 1.1 GEOGRAPHY OF THE MARSHALL ISLANDS. Approxi- Area mate No. (square miles) Latitude Longitude Name Type of Islets Lagoon Dry Land (Deg. Min.) (Deg. Min.) Marshall Islands 1,225 4,506.87 70.05 4 34' - 14 42' 160 47' - 172 10' Ratak Chain 641 1672.52 34.02 5 53' - 14 42' 168 53' - 172 10' 1 Bokak Atoll 10 30.13 1.25 14 32' - 14 42' 168 53' - 169 1' 2 Bikar Atoll 7 14.44 0.19 12 11' - 12 18' 170 4' - 170 9' 3 Utrok Atoll 10 22.29 0.94 11 12' - 11 12' 169 45' - 169 51' 4 Toke Atoll 6 35.96 0.22 11 5' - 11 30' 169 33' - 169 40' 5 Mejit Island 1 _ 0.72 10 16' - 10 18' 170 52' - 70 53' 6 Ailuk Atoll 55 68.47 2.07 10 12' - 10 27' 169 52' - 169 59' 7 Jemo Island 1 _ 0.06 10 7' 169 33' 8 Likiep Atoll 65 163.71 3.96 9 48' - 10 4' 168 58' - 169 20' 9 Wotje Atoll 75 241.06 3.16 9 21' - 9 32' 169 48' - 170 15' 10 Erkub Atoll 16 88.92 0.59 9 8' 170 0' 11 Maloelap Atoll 75 375.57 3.79 8 29' - 8 55' 170 50' - 171 15' 12 Aur Atoll 43 92.58 2.17 8 7' - 8 22' 171 1' - 171 12' 13 Majuro Atoll 64 113.92 3.75 7 3' - 7 13' 171 2' - 171 58' 14 Arno Atoll 103 130.77 5.00 6 58' - 7 18' 171 33' - 171 57' 15 Mili Atoll 92 294.70 6.15 5 53' - 6 16' 171 42' - 172 10' 16 Nadikdik Atoll 18 _ _ _ _ _ _ Ralik Chain 584 2834.35 36.03 4 34' - 11 42' 160 47' - 169 44' 17 Enewetak Atoll 44 387.99 2.26 11 20' - 11 41' 162 17' - 162 24' 18 Ujelang Atoll 30 25.47 0.67 9 46' - 9 53' 160 47' - 161 0' 19 Bikini Atoll 36 229.40 2.32 11 29' - 11 42' 165 12' - 165 34' 20 Rongdrik Atoll 14 55.38 0.65 11 15' - 11 25' 167 22' - 167 32' 21 Ronglap Atoll 61 387.77 3.07 11 9' - 11 -

Final Report Review of the Outer Islands Electrification Programmes in Rmi
FINAL REPORT REVIEW OF THE OUTER ISLANDS ELECTRIFICATION PROGRAMMES IN RMI RFP Reference: SPC 11/14 September 2012 Prepared by Global Sustainable Energy Solutions Pty. Limited For NORTH‐REP North Pacific ACP Renewable Energy and Energy Efficiency Project Acknowledgments This report was written by Global Sustainable energy Solutions Pty Ltd (GSES) for the North Pacific ACP Renewable Energy and Energy Efficiency Project (North REP). The authors wish to acknowledge the following people who provided information wither through meetings or via e‐mail communications in the preparation of the report: Mr Thomas Kijiner Jnr; Secretary of Resources & Development (At time of interview) Dr Hilda Heine, Minister for Education Mr Richard Bruce and Mr Kanchi Hosia Ministry of Education Ms Angeline Heine National Energy Planner, Energy Planning Division, Ministry of Resources & Development Mr Steve Wakefield Chief Technical Officer, Marshalls Energy Company Mr Billy Schultz Solar Engineer Marshalls Energy Company Mr Charles Abraham Mayor of Mejit: Ms Rufena Elooki : Mayor of Ailuk: Mr Walter Myazoe ; Assistant Energy Planner ,Energy Planning Division, Ministry of Resources & Development Ms Katerina Syngellakis, GIZ Energy Adviser Mr Rupeni Mario; Team Leader North REP Ms Arieta Gonelevu: North REP Energy Specialist (RMI) RFP Ref: 11/14 Review of the Outer Islands Electrification Programmes in RMI and FSM‐Final Report‐RMI i Contents Acknowledgments ................................................................................................................................... -

Together Through Life
Together Through Life An Ethnographic Study of Hierarchy, Equality, and Togetherness on the Epoon Atoll, Marshall Islands Ola Gunhildrud Berta Master’s thesis, the Department of Social Anthropology UNIVERSITY OF OSLO 2015 ii Together Through Life An Ethnographic Study of Hierarchy, Equality, and Togetherness on the Epoon Atoll, Marshall Islands Ola Gunhildrud Berta University of Oslo 2015 iii © Ola Gunhildrud Berta 2015 Together Through Life: An Ethnographic Study of Hierarchy, Equality, and Togetherness on the Epoon Atoll, Marshall Islands http://www.duo.uio.no/ Trykk: Reprosentralen, Universitetet i Oslo iv Abstract This thesis deals with everyday manifestations of hierarchy, equality and togetherness on the Epoon [Ebon] Atoll on the Marshall Islands. More particularly, it investigates some of the ways in which people on Epoon evoke and invoke hierarchy and egalitarianism, depending on context. People on Epoon have a striking tendency to stress equality and togetherness among themselves, in public speeches as well as in their cooking hut conversations. This emphasis on egalitarian values is visible in most social dynamics on the atoll. Interestingly, these egalitarian ideals often stand in direct opposition to a hierarchical form of social structure. Anthropologists have typically described the Marshallese social structure as a class-based hierarchy, ranging from commoners to high chiefs, with hierarchical differences within each class. Having some forms of hierarchical organization, however, does not necessarily mean inequality regarding social life on Epoon. On the contrary, I argue that, even with the presence of hierarchy, equality stands out as an encompassing cultural value. Through four ethnographic chapters, I seek to illustrate some of the ways in which this plays out in daily life.