Modular Effects of Gene Promoters and Chromatin Environments on Noise in Gene Expression
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Exploring Autophagy with Gene Ontology
Autophagy ISSN: 1554-8627 (Print) 1554-8635 (Online) Journal homepage: https://www.tandfonline.com/loi/kaup20 Exploring autophagy with Gene Ontology Paul Denny, Marc Feuermann, David P. Hill, Ruth C. Lovering, Helene Plun- Favreau & Paola Roncaglia To cite this article: Paul Denny, Marc Feuermann, David P. Hill, Ruth C. Lovering, Helene Plun- Favreau & Paola Roncaglia (2018) Exploring autophagy with Gene Ontology, Autophagy, 14:3, 419-436, DOI: 10.1080/15548627.2017.1415189 To link to this article: https://doi.org/10.1080/15548627.2017.1415189 © 2018 The Author(s). Published by Informa UK Limited, trading as Taylor & Francis Group. View supplementary material Published online: 17 Feb 2018. Submit your article to this journal Article views: 1097 View Crossmark data Full Terms & Conditions of access and use can be found at https://www.tandfonline.com/action/journalInformation?journalCode=kaup20 AUTOPHAGY, 2018 VOL. 14, NO. 3, 419–436 https://doi.org/10.1080/15548627.2017.1415189 RESEARCH PAPER - BASIC SCIENCE Exploring autophagy with Gene Ontology Paul Denny a,†,§, Marc Feuermann b,§, David P. Hill c,f,§, Ruth C. Lovering a,§, Helene Plun-Favreau d and Paola Roncaglia e,f,§ aFunctional Gene Annotation, Institute of Cardiovascular Science, University College London, London, UK; bSIB Swiss Institute of Bioinformatics, Geneva, Switzerland; cThe Jackson Laboratory, Bar Harbor, ME, USA; dDepartment of Molecular Neuroscience, UCL Institute of Neurology, London, UK; eEuropean Bioinformatics Institute (EMBL-EBI), European Molecular Biology Laboratory, Wellcome Genome Campus, Hinxton, Cambridge, UK; fThe Gene Ontology Consortium ABSTRACT ARTICLE HISTORY Autophagy is a fundamental cellular process that is well conserved among eukaryotes. It is one of the Received 18 May 2017 strategies that cells use to catabolize substances in a controlled way. -

Detailed Characterization of Human Induced Pluripotent Stem Cells Manufactured for Therapeutic Applications
Stem Cell Rev and Rep DOI 10.1007/s12015-016-9662-8 Detailed Characterization of Human Induced Pluripotent Stem Cells Manufactured for Therapeutic Applications Behnam Ahmadian Baghbaderani 1 & Adhikarla Syama2 & Renuka Sivapatham3 & Ying Pei4 & Odity Mukherjee2 & Thomas Fellner1 & Xianmin Zeng3,4 & Mahendra S. Rao5,6 # The Author(s) 2016. This article is published with open access at Springerlink.com Abstract We have recently described manufacturing of hu- help determine which set of tests will be most useful in mon- man induced pluripotent stem cells (iPSC) master cell banks itoring the cells and establishing criteria for discarding a line. (MCB) generated by a clinically compliant process using cord blood as a starting material (Baghbaderani et al. in Stem Cell Keywords Induced pluripotent stem cells . Embryonic stem Reports, 5(4), 647–659, 2015). In this manuscript, we de- cells . Manufacturing . cGMP . Consent . Markers scribe the detailed characterization of the two iPSC clones generated using this process, including whole genome se- quencing (WGS), microarray, and comparative genomic hy- Introduction bridization (aCGH) single nucleotide polymorphism (SNP) analysis. We compare their profiles with a proposed calibra- Induced pluripotent stem cells (iPSCs) are akin to embryonic tion material and with a reporter subclone and lines made by a stem cells (ESC) [2] in their developmental potential, but dif- similar process from different donors. We believe that iPSCs fer from ESC in the starting cell used and the requirement of a are likely to be used to make multiple clinical products. We set of proteins to induce pluripotency [3]. Although function- further believe that the lines used as input material will be used ally identical, iPSCs may differ from ESC in subtle ways, at different sites and, given their immortal status, will be used including in their epigenetic profile, exposure to the environ- for many years or even decades. -

Exploring Extracellular Vesicles Biogenesis in Hypothalamic Cells Through a Heavy Isotope Pulse/Trace Proteomic Approach
cells Article Exploring Extracellular Vesicles Biogenesis in Hypothalamic Cells through a Heavy Isotope Pulse/Trace Proteomic Approach Chee Fan Tan 1,2 , Hui San Teo 2, Jung Eun Park 2, Bamaprasad Dutta 2, Shun Wilford Tse 2, Melvin Khee-Shing Leow 3,4,5 , Walter Wahli 3,6 and Siu Kwan Sze 2,* 1 NTU Institute for Health Technologies, Interdisciplinary Graduate School, Nanyang Technological University, Singapore 637335, Singapore; [email protected] 2 School of Biological Sciences, Nanyang Technological University, Singapore 637551, Singapore; [email protected] (H.S.T.); [email protected] (J.E.P.); [email protected] (B.D.); [email protected] (S.W.T.) 3 Lee Kong Chian School of Medicine, Nanyang Technological University, Singapore 636921, Singapore; [email protected] (M.K.-S.L.); [email protected] (W.W.) 4 Department of Endocrinology, Tan Tock Seng Hospital, Singapore 308433, Singapore 5 Cardiovascular and Metabolic Disorder Program, Duke-NUS Medical School, Singapore 169857, Singapore 6 Center for Integrative Genomics, University of Lausanne, Le Génopode, CH-1015 Lausanne, Switzerland * Correspondence: [email protected]; Tel.: +65-6514-1006; Fax: +65-6791-3856 Received: 24 March 2020; Accepted: 21 May 2020; Published: 25 May 2020 Abstract: Studies have shown that the process of extracellular vesicles (EVs) secretion and lysosome status are linked. When the lysosome is under stress, the cells would secrete more EVs to maintain cellular homeostasis. However, the process that governs lysosomal activity and EVs secretion remains poorly defined and we postulated that certain proteins essential for EVs biogenesis are constantly synthesized and preferentially sorted to the EVs rather than the lysosome. -

Changes in Prostate Gene Expression in Men Undergoing an Intensive Nutrition and Lifestyle Intervention
Changes in prostate gene expression in men undergoing an intensive nutrition and lifestyle intervention Dean Ornish*†‡, Mark Jesus M. Magbanua§, Gerdi Weidner*, Vivian Weinberg¶, Colleen Kemp*, Christopher Green§, Michael D. Mattie§, Ruth Marlin*, Jeff Simkoʈ, Katsuto Shinohara§, Christopher M. Haqq§ and Peter R. Carroll§ §Department of Urology, The Helen Diller Family Comprehensive Cancer Center, and ʈDepartment of Pathology, University of California, 2340 Sutter Street, San Francisco, CA 94115; *Preventive Medicine Research Institute, 900 Bridgeway, Sausalito, CA 94965; †Department of Medicine, School of Medicine, University of California, 505 Parnassus Avenue, San Francisco, CA 94143; and ¶Biostatistics Core, The Helen Diller Family Comprehensive Cancer Center, University of California, 513 Parnassus Avenue, Box 0127, San Francisco, CA 94143 Communicated by J. Craig Venter, The J. Craig Venter Institute, Rockville, MD, April 2, 2008 (received for review February 13, 2008) Epidemiological and prospective studies indicate that comprehensive indolent low-risk prostate cancers, defined by strict clinical and lifestyle changes may modify the progression of prostate cancer. pathologic criteria designed to minimize the risk for metastatic However, the molecular mechanisms by which improvements in diet disease as a result of study participation (9). The 30 men who and lifestyle might affect the prostate microenvironment are poorly enrolled did not undergo surgery or radiation therapy to treat their understood. We conducted a pilot study to examine changes in low-risk tumors; rather, they underwent comprehensive lifestyle prostate gene expression in a unique population of men with low-risk changes (low-fat, whole-foods, plant-based nutrition; stress man- prostate cancer who declined immediate surgery, hormonal therapy, agement techniques; moderate exercise; and participation in a or radiation and participated in an intensive nutrition and lifestyle psychosocial group support). -

Downloaded Per Proteome Cohort Via the Web- Site Links of Table 1, Also Providing Information on the Deposited Spectral Datasets
www.nature.com/scientificreports OPEN Assessment of a complete and classifed platelet proteome from genome‑wide transcripts of human platelets and megakaryocytes covering platelet functions Jingnan Huang1,2*, Frauke Swieringa1,2,9, Fiorella A. Solari2,9, Isabella Provenzale1, Luigi Grassi3, Ilaria De Simone1, Constance C. F. M. J. Baaten1,4, Rachel Cavill5, Albert Sickmann2,6,7,9, Mattia Frontini3,8,9 & Johan W. M. Heemskerk1,9* Novel platelet and megakaryocyte transcriptome analysis allows prediction of the full or theoretical proteome of a representative human platelet. Here, we integrated the established platelet proteomes from six cohorts of healthy subjects, encompassing 5.2 k proteins, with two novel genome‑wide transcriptomes (57.8 k mRNAs). For 14.8 k protein‑coding transcripts, we assigned the proteins to 21 UniProt‑based classes, based on their preferential intracellular localization and presumed function. This classifed transcriptome‑proteome profle of platelets revealed: (i) Absence of 37.2 k genome‑ wide transcripts. (ii) High quantitative similarity of platelet and megakaryocyte transcriptomes (R = 0.75) for 14.8 k protein‑coding genes, but not for 3.8 k RNA genes or 1.9 k pseudogenes (R = 0.43–0.54), suggesting redistribution of mRNAs upon platelet shedding from megakaryocytes. (iii) Copy numbers of 3.5 k proteins that were restricted in size by the corresponding transcript levels (iv) Near complete coverage of identifed proteins in the relevant transcriptome (log2fpkm > 0.20) except for plasma‑derived secretory proteins, pointing to adhesion and uptake of such proteins. (v) Underrepresentation in the identifed proteome of nuclear‑related, membrane and signaling proteins, as well proteins with low‑level transcripts. -

Content Based Search in Gene Expression Databases and a Meta-Analysis of Host Responses to Infection
Content Based Search in Gene Expression Databases and a Meta-analysis of Host Responses to Infection A Thesis Submitted to the Faculty of Drexel University by Francis X. Bell in partial fulfillment of the requirements for the degree of Doctor of Philosophy November 2015 c Copyright 2015 Francis X. Bell. All Rights Reserved. ii Acknowledgments I would like to acknowledge and thank my advisor, Dr. Ahmet Sacan. Without his advice, support, and patience I would not have been able to accomplish all that I have. I would also like to thank my committee members and the Biomed Faculty that have guided me. I would like to give a special thanks for the members of the bioinformatics lab, in particular the members of the Sacan lab: Rehman Qureshi, Daisy Heng Yang, April Chunyu Zhao, and Yiqian Zhou. Thank you for creating a pleasant and friendly environment in the lab. I give the members of my family my sincerest gratitude for all that they have done for me. I cannot begin to repay my parents for their sacrifices. I am eternally grateful for everything they have done. The support of my sisters and their encouragement gave me the strength to persevere to the end. iii Table of Contents LIST OF TABLES.......................................................................... vii LIST OF FIGURES ........................................................................ xiv ABSTRACT ................................................................................ xvii 1. A BRIEF INTRODUCTION TO GENE EXPRESSION............................. 1 1.1 Central Dogma of Molecular Biology........................................... 1 1.1.1 Basic Transfers .......................................................... 1 1.1.2 Uncommon Transfers ................................................... 3 1.2 Gene Expression ................................................................. 4 1.2.1 Estimating Gene Expression ............................................ 4 1.2.2 DNA Microarrays ...................................................... -

Table S1 Genes with Coefficients of Variation (CV) Ratios (CVGHS/CVSD) in the Upper and Lower 10% of for Expression in Genetic Hypercalciuric Stone-Forming (GHS) And
Table S1 Genes with coefficients of variation (CV) ratios (CVGHS/CVSD) in the upper and lower 10% of for expression in Genetic Hypercalciuric Stone-forminG (GHS) and Sprague-Dawley (SD) rats, assayed usinG Affymetrix Rat Genome 230 GeneChips. The location of the microsatellite or quantitative trait locus (QTL) position (Loc) in base pairs (bp), gene initiation base pair (BP), chromosomal banding position and RGD or GenBank name are given.Rat Genome Database (RGD) (www.rgd.mcw.edu), GenBank (http://www.ncbi.nlm.nih.gov/Genbank/) and Entrez (http://www.ncbi.nlm.nih.gov/sites/entrez). Variance in gene expression was ranked according to CVGHS/CVSD ratio for all renal (8846 genes, Pctl, Renal) and duodenal genes (9038 genes, Pctl, Duod). Gene Pctl, Pctl, QTL Gene (GenBank) Renal Duod BP GenBank/RGD Description D1Mit95 Riok2 (BG371773) 91.3 52.4 55,897,246 RIO kinase 2 66,832,211 Tmc4 (BF545988) - 93.6 63,582,715 transmembrane channel-like gene family 4 Isoc2b (BM385414) 2.3 - 67,689,183 Isochorismatase domain containing 2b Suv420h2 (AW525235) 69.9 97.3 67,815,980 suppressor of variegation 4-20 homolog 2 (Drosophila) Lilrb4 (BF282961) 95.5 - 69,165,497 leukocyte immunoglobulin-like receptor, subfamily B, member 4 Zbtb45 (BI289556) - 10.0 72,912,203 zinc finger and BTB domain containing 45 Sepw1 (NM_013027) 7.5 - 76,249,869 selenoprotein W, muscle 1 Chmp2a (AW434104) 0.1 86.9 72,889,348 chromatin modifying protein 2a Tmem160 (AWS25031) 97.6 2.2 76,716,254 transmembrane protein Pnmal2 (BI282311) 99.6 - 77,277,341 PNMA-like 2 Dmpk (AI044427) 98.9 7.3 78,449,323 dystrophia myotonica-protein kinase; serine-threonine kinase Vasp (AW520792) 99.3 - 78,621,478 vasodilator-stimulated phosphoprotein Ercc1 (AA892791) 90.6 - 78,711,248 excision repair cross-complementing rodent repair deficiency, complementation G. -
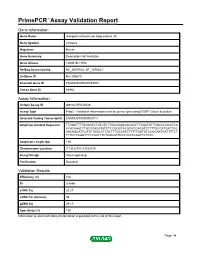
Primepcr™Assay Validation Report
PrimePCR™Assay Validation Report Gene Information Gene Name charged multivesicular body protein 2A Gene Symbol Chmp2a Organism Mouse Gene Summary Description Not Available Gene Aliases 1500016L11Rik RefSeq Accession No. NC_000073.6, NT_187034.1 UniGene ID Mm.295670 Ensembl Gene ID ENSMUSG00000033916 Entrez Gene ID 68953 Assay Information Unique Assay ID qMmuCIP0034548 Assay Type Probe - Validation information is for the primer pair using SYBR® Green detection Detected Coding Transcript(s) ENSMUST00000005711 Amplicon Context Sequence TTGGATTTTAGAGTCTGTATCTTGAGGGACACAGCTTGGATGTTGGCCCGCATCA ACACAAACTTGCGTACATATCTCCGGGTACGCACCAGGTCTTTTGCCATGATTCG AACAGCATCCATCTGGCCTTGCTTTGCCATCTTTTTGATGTCCGCAATGATTTTCT TTTCCTGGGTTTCTAGTTTCTGTCGTTCCCTGTCCAGTTCTCTC Amplicon Length (bp) 180 Chromosome Location 7:13032703-13033919 Assay Design Intron-spanning Purification Desalted Validation Results Efficiency (%) 100 R2 0.9999 cDNA Cq 20.27 cDNA Tm (Celsius) 85 gDNA Cq 39.13 Specificity (%) 100 Information to assist with data interpretation is provided at the end of this report. Page 1/4 PrimePCR™Assay Validation Report Chmp2a, Mouse Amplification Plot Amplification of cDNA generated from 25 ng of universal reference RNA Melt Peak Melt curve analysis of above amplification Standard Curve Standard curve generated using 20 million copies of template diluted 10-fold to 20 copies Page 2/4 PrimePCR™Assay Validation Report Products used to generate validation data Real-Time PCR Instrument CFX384 Real-Time PCR Detection System Reverse Transcription Reagent iScript™ Advanced cDNA Synthesis Kit for RT-qPCR Real-Time PCR Supermix SsoAdvanced™ SYBR® Green Supermix Experimental Sample qPCR Mouse Reference Total RNA Data Interpretation Unique Assay ID This is a unique identifier that can be used to identify the assay in the literature and online. Detected Coding Transcript(s) This is a list of the Ensembl transcript ID(s) that this assay will detect. -

The ESCRT-III Proteins IST1 and CHMP1B Assemble Around Nucleic Acids
bioRxiv preprint doi: https://doi.org/10.1101/386532; this version posted August 7, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. The ESCRT-III proteins IST1 and CHMP1B assemble around nucleic acids Nathaniel Talledge#1,2,3,4, John McCullough#1, Dawn Wenzel1, Henry C. Nguyen2,3, Matthew S. Lalonde1, Monika Bajorek1, Jack Skalicky1, Adam Frost1,2,3,4,*, Wesley I. Sundqust1,* 1 Department of Biochemistry, University of Utah, Salt Lake City, UT 84112, United States. 2 Department of Biochemistry & Biophysics, University of California, San Francisco, San Francisco, CA 94158, United States. 3 California Institute for Quantitative Biomedical Research, University of California, San Francisco, United States. 4 Chan Zuckerberg Biohub, United States. # These authors contributed equally *Co-corresponding authors 1 bioRxiv preprint doi: https://doi.org/10.1101/386532; this version posted August 7, 2018. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted bioRxiv a license to display the preprint in perpetuity. It is made available under aCC-BY-NC-ND 4.0 International license. ABSTRACT ESCRT-III proteins can promote inside-out or outside-in membrane tubulation and fission. In addition, several observations suggest that ESCRT factors may also associate with nucleic acids during development, different stages of the cell cycle, and during retro- transposition of parasitic nucleic acids like LINE1 elements. -

Coexpression Networks Based on Natural Variation in Human Gene Expression at Baseline and Under Stress
University of Pennsylvania ScholarlyCommons Publicly Accessible Penn Dissertations Fall 2010 Coexpression Networks Based on Natural Variation in Human Gene Expression at Baseline and Under Stress Renuka Nayak University of Pennsylvania, [email protected] Follow this and additional works at: https://repository.upenn.edu/edissertations Part of the Computational Biology Commons, and the Genomics Commons Recommended Citation Nayak, Renuka, "Coexpression Networks Based on Natural Variation in Human Gene Expression at Baseline and Under Stress" (2010). Publicly Accessible Penn Dissertations. 1559. https://repository.upenn.edu/edissertations/1559 This paper is posted at ScholarlyCommons. https://repository.upenn.edu/edissertations/1559 For more information, please contact [email protected]. Coexpression Networks Based on Natural Variation in Human Gene Expression at Baseline and Under Stress Abstract Genes interact in networks to orchestrate cellular processes. Here, we used coexpression networks based on natural variation in gene expression to study the functions and interactions of human genes. We asked how these networks change in response to stress. First, we studied human coexpression networks at baseline. We constructed networks by identifying correlations in expression levels of 8.9 million gene pairs in immortalized B cells from 295 individuals comprising three independent samples. The resulting networks allowed us to infer interactions between biological processes. We used the network to predict the functions of poorly-characterized human genes, and provided some experimental support. Examining genes implicated in disease, we found that IFIH1, a diabetes susceptibility gene, interacts with YES1, which affects glucose transport. Genes predisposing to the same diseases are clustered non-randomly in the network, suggesting that the network may be used to identify candidate genes that influence disease susceptibility. -

Table S1. 103 Ferroptosis-Related Genes Retrieved from the Genecards
Table S1. 103 ferroptosis-related genes retrieved from the GeneCards. Gene Symbol Description Category GPX4 Glutathione Peroxidase 4 Protein Coding AIFM2 Apoptosis Inducing Factor Mitochondria Associated 2 Protein Coding TP53 Tumor Protein P53 Protein Coding ACSL4 Acyl-CoA Synthetase Long Chain Family Member 4 Protein Coding SLC7A11 Solute Carrier Family 7 Member 11 Protein Coding VDAC2 Voltage Dependent Anion Channel 2 Protein Coding VDAC3 Voltage Dependent Anion Channel 3 Protein Coding ATG5 Autophagy Related 5 Protein Coding ATG7 Autophagy Related 7 Protein Coding NCOA4 Nuclear Receptor Coactivator 4 Protein Coding HMOX1 Heme Oxygenase 1 Protein Coding SLC3A2 Solute Carrier Family 3 Member 2 Protein Coding ALOX15 Arachidonate 15-Lipoxygenase Protein Coding BECN1 Beclin 1 Protein Coding PRKAA1 Protein Kinase AMP-Activated Catalytic Subunit Alpha 1 Protein Coding SAT1 Spermidine/Spermine N1-Acetyltransferase 1 Protein Coding NF2 Neurofibromin 2 Protein Coding YAP1 Yes1 Associated Transcriptional Regulator Protein Coding FTH1 Ferritin Heavy Chain 1 Protein Coding TF Transferrin Protein Coding TFRC Transferrin Receptor Protein Coding FTL Ferritin Light Chain Protein Coding CYBB Cytochrome B-245 Beta Chain Protein Coding GSS Glutathione Synthetase Protein Coding CP Ceruloplasmin Protein Coding PRNP Prion Protein Protein Coding SLC11A2 Solute Carrier Family 11 Member 2 Protein Coding SLC40A1 Solute Carrier Family 40 Member 1 Protein Coding STEAP3 STEAP3 Metalloreductase Protein Coding ACSL1 Acyl-CoA Synthetase Long Chain Family Member 1 Protein -

Proteomic Analysis Uncovers Measles Virus Protein C
bioRxiv preprint doi: https://doi.org/10.1101/2020.05.08.084418; this version posted May 9, 2020. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Proteomic Analysis Uncovers Measles Virus Protein C Interaction with p65/iASPP/p53 Protein Complex Alice Meignié1,2*, Chantal Combredet1*, Marc Santolini 3,4, István A. Kovács4,5,6, Thibaut Douché7, Quentin Giai Gianetto 7,8, Hyeju Eun9, Mariette Matondo7, Yves Jacob10, Regis Grailhe9, Frédéric Tangy1**, and Anastassia V. Komarova1, 10** 1 Viral Genomics and Vaccination Unit, Department of Virology, Institut Pasteur, CNRS UMR-3569, 75015 Paris, France 2 Université Paris Diderot, Sorbonne Paris Cité, Paris, France 3 Center for Research and Interdisciplinarity (CRI), Université de Paris, INSERM U1284 4 Network Science Institute and Department of Physics, Northeastern University, Boston, MA 02115, USA 5 Department of Physics and Astronomy, Northwestern University, Evanston, IL 60208-3109, USA 6 Department of Network and Data Science, Central European University, Budapest, H-1051, Hungary 7 Proteomics platform, Mass Spectrometry for Biology Unit (MSBio), Institut Pasteur, CNRS USR 2000, Paris, France. 8 Bioinformatics and Biostatistics Hub, Computational Biology Department, Institut Pasteur, CNRS USR3756, Paris, France 9 Technology Development Platform, Institut Pasteur Korea, Seongnam-si, Republic of Korea 10 Laboratory of Molecular Genetics of RNA Viruses, Institut Pasteur, CNRS UMR-3569,