Silurian Spathacalymene Tillman, 1960 (Trilobita) from Poland
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

The Case of the Diminutive Trilobite Flexicalymene Retrorsa Minuens from the Cincinnatian Series (Upper Ordovician), Cincinnati Region
EVOLUTION & DEVELOPMENT 9:5, 483–498 (2007) Evaluating paedomorphic heterochrony in trilobites: the case of the diminutive trilobite Flexicalymene retrorsa minuens from the Cincinnatian Series (Upper Ordovician), Cincinnati region Brenda R. Hundaa,Ã and Nigel C. Hughesb aCincinnati Museum Center, 1301 Western Avenue, Cincinnati, OH 45203, USA bDepartment of Earth Sciences, University of California, Riverside, CA 92521, USA ÃAuthor for correspondence (email: [email protected]) SUMMARY Flexicalymene retrorsa minuens from the upper- rate of progress along a common ontogenetic trajectory with most 3 m of the Waynesville Formation of the Cincinnatian respect to size, coupled with growth cessation at a small size, Series (Upper Ordovician) of North America lived ‘‘sequential’’ progenesis, or non-uniform changes in the rate of approximately 445 Ma and exhibited marked reduction in progress along a shared ontogenetic trajectory with respect to maximum size relative to its stratigraphically subjacent sister size, can also be rejected. Rather, differences between these subspecies, Flexicalymene retrorsa retrorsa. Phylogenetic subspecies are more consistent with localized changes in analysis is consistent with the notion that F. retrorsa retrorsa rates of character development than with a global hetero- was the ancestor of F. retrorsa minuens. F. retrorsa minuens chronic modification of the ancestral ontogeny. The evolution has been claimed to differ from F. retrorsa retrorsa ‘‘in size of F. retrorsa minuens from F. retrorsa retrorsa was largely alone,’’ and thus presents a plausible example of global dominated by modifications of the development of characters paedomorphic evolution in trilobites. Despite strong similarity already evident in the ancestral ontogeny, not by the origin of in the overall form of the two subspecies, F. -

United States
DEPARTMENT OF THE INTERIOR BULLETIN OF THE UNITED STATES ISTo. 146 WASHINGTON GOVERNMENT Pit IN TING OFFICE 189C UNITED STATES GEOLOGICAL SURVEY CHAKLES D. WALCOTT, DI11ECTOK BIBLIOGRAPHY AND INDEX NORTH AMEEICAN GEOLOGY, PALEONTOLOGY, PETEOLOGT, AND MINERALOGY THE YEA.R 1895 FEED BOUGHTON WEEKS WASHINGTON Cr O V E U N M K N T P K 1 N T I N G OFFICE 1890 CONTENTS. Page. Letter of trail smittal...... ....................... .......................... 7 Introduction.............'................................................... 9 List of publications examined............................................... 11 Classified key to tlio index .......................................... ........ 15 Bibliography ............................................................... 21 Index....................................................................... 89 LETTER OF TRANSMITTAL DEPARTMENT OF THE INTEEIOE, UNITED STATES GEOLOGICAL SURVEY, DIVISION OF GEOLOGY, Washington, D. 0., June 23, 1896. SIR: I have the honor to transmit herewith the manuscript of a Bibliography and Index of North American Geology, Paleontology, Petrology, and Mineralogy for the year 1895, and to request that it be published as a bulletin of the Survey. Very respectfully, F. B. WEEKS. Hon. CHARLES D. WALCOTT, Director United States Geological Survey. 1 BIBLIOGRAPHY AND INDEX OF NORTH AMERICAN GEOLOGY, PALEONTOLOGY, PETROLOGY, AND MINER ALOGY FOR THE YEAR 1895. By FRED BOUGHTON WEEKS. INTRODUCTION. The present work comprises a record of publications on North Ameri can geology, paleontology, petrology, and mineralogy for the year 1895. It is planned on the same lines as the previous bulletins (Nos. 130 and 135), excepting that abstracts appearing in regular periodicals have been omitted in this volume. Bibliography. The bibliography consists of full titles of separate papers, classified by authors, an abbreviated reference to the publica tion in which the paper is printed, and a brief summary of the con tents, each paper being numbered for index reference. -

An Inventory of Trilobites from National Park Service Areas
Sullivan, R.M. and Lucas, S.G., eds., 2016, Fossil Record 5. New Mexico Museum of Natural History and Science Bulletin 74. 179 AN INVENTORY OF TRILOBITES FROM NATIONAL PARK SERVICE AREAS MEGAN R. NORR¹, VINCENT L. SANTUCCI1 and JUSTIN S. TWEET2 1National Park Service. 1201 Eye Street NW, Washington, D.C. 20005; -email: [email protected]; 2Tweet Paleo-Consulting. 9149 79th St. S. Cottage Grove. MN 55016; Abstract—Trilobites represent an extinct group of Paleozoic marine invertebrate fossils that have great scientific interest and public appeal. Trilobites exhibit wide taxonomic diversity and are contained within nine orders of the Class Trilobita. A wealth of scientific literature exists regarding trilobites, their morphology, biostratigraphy, indicators of paleoenvironments, behavior, and other research themes. An inventory of National Park Service areas reveals that fossilized remains of trilobites are documented from within at least 33 NPS units, including Death Valley National Park, Grand Canyon National Park, Yellowstone National Park, and Yukon-Charley Rivers National Preserve. More than 120 trilobite hototype specimens are known from National Park Service areas. INTRODUCTION Of the 262 National Park Service areas identified with paleontological resources, 33 of those units have documented trilobite fossils (Fig. 1). More than 120 holotype specimens of trilobites have been found within National Park Service (NPS) units. Once thriving during the Paleozoic Era (between ~520 and 250 million years ago) and becoming extinct at the end of the Permian Period, trilobites were prone to fossilization due to their hard exoskeletons and the sedimentary marine environments they inhabited. While parks such as Death Valley National Park and Yukon-Charley Rivers National Preserve have reported a great abundance of fossilized trilobites, many other national parks also contain a diverse trilobite fauna. -

Late Silurian Trilobite Palaeobiology And
LATE SILURIAN TRILOBITE PALAEOBIOLOGY AND BIODIVERSITY by ANDREW JAMES STOREY A thesis submitted to the University of Birmingham for the degree of DOCTOR OF PHILOSOPHY School of Geography, Earth and Environmental Sciences University of Birmingham February 2012 University of Birmingham Research Archive e-theses repository This unpublished thesis/dissertation is copyright of the author and/or third parties. The intellectual property rights of the author or third parties in respect of this work are as defined by The Copyright Designs and Patents Act 1988 or as modified by any successor legislation. Any use made of information contained in this thesis/dissertation must be in accordance with that legislation and must be properly acknowledged. Further distribution or reproduction in any format is prohibited without the permission of the copyright holder. ABSTRACT Trilobites from the Ludlow and Přídolí of England and Wales are described. A total of 15 families; 36 genera and 53 species are documented herein, including a new genus and seventeen new species; fourteen of which remain under open nomenclature. Most of the trilobites in the British late Silurian are restricted to the shelf, and predominantly occur in the Elton, Bringewood, Leintwardine, and Whitcliffe groups of Wales and the Welsh Borderland. The Elton to Whitcliffe groups represent a shallowing upwards sequence overall; each is characterised by a distinct lithofacies and fauna. The trilobites and brachiopods of the Coldwell Formation of the Lake District Basin are documented, and are comparable with faunas in the Swedish Colonus Shale and the Mottled Mudstones of North Wales. Ludlow trilobite associations, containing commonly co-occurring trilobite taxa, are defined for each palaeoenvironment. -

North American Geology, Paleontology, Petrology, and Mineralogy
Bulletin No. 240 Series G, Miscellaneous, 28 DEPARTMENT OF THE INTERIOR UNITED STATES GEOLOGICAL SURVEY CHARLES D. VVALCOTT, DIRECTOR BIBIIOGRAP.HY AND INDEX OF NORTH AMERICAN GEOLOGY, PALEONTOLOGY, PETROLOGY, AND MINERALOGY FOR THE YEAJR, 19O3 BY IFIRIEID WASHINGTON GOVERNMENT PRINTING OFFICE 1904 CONTENTS Page. Letter of transmittal...................................................... 5 Introduction.....:....................................,.................. 7 List of publications examined ............................................. 9 Bibliography............................................................. 13 Addenda to bibliographies J'or previous years............................... 139 Classi (led key to the index................................................ 141 Index .._.........;.................................................... 149 LETTER OF TRANSMITTAL DEPARTMENT OF THE INTERIOR, UNITED STATES GEOLOGICAL SURVEY, Washington, D. 0. , June 7, 1904.. SIR: I have the honor to transmit herewith the manuscript of a bibliography and index of North American geology, paleontology, petrology, and mineralogy for the year 1903, and to request that it be published as a bulletin of the Survey. Very respectfully, F. B. WEEKS, Libraria/ii. Hon. CHARLES D. WALCOTT, Director United States Geological Survey. BIBLIOGRAPHY AND INDEX OF NORTH AMERICAN GEOLOGY,- PALEONTOLOGY, PETROLOGY, AND MINERALOGY FOR THE YEAR 1903. By FRED BOUGHTON WEEKS. INTRODUCTION, The arrangement of the material of the Bibliography and Index f Or 1903 is similar -

Iii Silurian Fauna
III SILURIAN FAUNA By AUGUST F. FOERSTE THE SILURIAN FAUNA OF KENTUCKY By AUGUST F. FOERSTE THE SOURCE AND AFFINITIES OF THE SILURIAN FAUNAS In prehistoric times the North American continent was invaded by oceanic faunas from various directions. The tilting of the continent toward the south permitted the invasion of oceanic faunas from the Gulf of Mexico, and tilting toward the east, permitted invasions from the North Atlantic. There have been invasions also from the Arctic and Pacific. Some of these invasions extended far inland. Some from the Gulf of Mexico spread northward along the Mississippi embayment as far as the southern part of Canada, and some from the North Atlantic reached Ohio and Kentucky. As far as known, none of the invasions from the Arctic or Pacific reached Kentucky. In most Silurian faunas, the brachiopod element predominates. Three of the southern invasions—the Brassfield, Osgood, and Waldron—are characterized by the abundance of bryozoans, in addition to the brachiopods. The Brassfield and Waldron contain also a considerable number of gastropods, pelecypods, cephalopods, and trilobites, all of which are relatively scarce in the Osgood and Louisville. In the Osgood, on the contrary, there are numerous cystids, though almost all of these were described originally under the single generic name Holocystites. The Waldron is characterized by the considerable variety of crinoids. In the Laurel and Louisville bryozoans are rare. The Laurel is now characterized by numerous species of crinoids, a considerable number of which show affinities with crinoids occurring in Gotland and England. Possibly these invaded the American continent from some North Atlantic source. -

Legacy of the Locust—Dudley and Its Famous Trilobite Calymene Blumenbachii
LEGACY OF THE LOCUST—DUDLEY AND ITS FAMOUS TRILOBITE CALYMENE BLUMENBACHII DONALD G. MIKULIC AND JOANNE KLUESSENDORF Illinois State Geological Survey, Champaign, IL 61820, [email protected], and Weis Earth Science Museum, University of Wisconsin-Fox Valley, Menasha, Wisconsin 54952, [email protected] ABSTRACT—The trilobite Calymene blumenbachii from the Silurian at Dudley, England, had a fundamental role in the early study of this prominent group of extinct arthropods. Discovered during the mid-1700s, this was the first trilo- bite known from numerous complete and well-preserved fossils anywhere in the world. Commonly known as the Dudley Fossil or Dudley Locust, exceptional specimens of this trilobite became widely distributed in collections throughout Europe. As a result, they were central to the most influential trilobite papers of the time including those of Walch (1771) and Brongniart (1822). Many basic characteristics of the group, including their ability to enroll, were first established through the study of these fossils. In turn, this information provided the key evidence used to estab- lish the arthropod affinities of this group. During the late eighteenth century, all trilobites were commonly referred to as Dudley Fossils, and demonstrate the initial importance and prominence of this species. It became the standard of comparison in trilobite research, as well as the textbook example for these fossils. No other trilobite contributed as much to the early understanding of these ancient animals. The scientific prominence of Calymene blumenbachii derived from geologic and economic factors. The limestones at Dudley contained an exceptionally rich biota of well-preserved Silurian fossils, of which this trilobite was the most notable. -
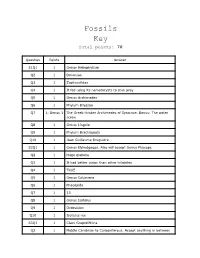
Fossils Key Total Points: 70
Fossils Key Total points: 70 Question Points Answer S1Q1 1 Genus Heliophyllum Q2 1 Devonian Q3 1 Zaphrentidae Q4 1 It fed using its nematocysts to stun prey Q5 1 Genus Archimedes Q6 1 Phylum Bryozoa Q7 1; Bonus 1 The Greek thinker Archimedes of Syracuse. Bonus: The water screw Q8 1 Genus Lingula Q9 1 Phylum Brachiopoda Q10 1 Jean Guillaume Bruguière S2Q1 1 Genus Eldredgeops. Also will accept Genus Phacops Q2 1 Huge glabella Q3 1 It had better vision than other trilobites Q4 1 TRUE Q5 1 Genus Calymene Q6 1 Phacopida Q7 1 13 Q8 1 Genus Isotelus Q9 1 Ordovician Q10 1 Isotelus rex S3Q1 1 Class Graptolithina Q2 1 Middle Cambrian to Carboniferous. Accept anything in between Q3 1 Accept graptos or lithos Q4 1 Phylum Hemichordata Q5 1 Genus Gryphaea Q6 1 Devil’s toenails Q7 1 TRUE Q8 1 Genus Dactylioceras Q9 1 Subclass Ammonoidea (Ammonoids) Q10 1 dactyl, meaning finger S4Q1 1 Genus Bothriolepis Q2 1 Bothriolepididae Q3 1 That external nasal openings lay on the dorsal side of the head near the eyes Q4 1 They break down easily and decompose much faster than hard tissues Q5 1 Genus Acanthostega Q6 1 Recognizable limbs Q7 1 TRUE Q8 1 Order Coelacanthiformes Q9 1 Class Sarcopterygii Q10 1 TRUE S5Q1 1 Genus Diplodocus Q2 1 1877 Q3 1 Othniel Charles Marsh Q4 1 Its forelimbs are slightly shorter than its hind limbs Q5 1 Genus Tyrannosaurus Q6 1 Order Saurischia Q7 1 Western North America. Will also accept North America Q8 1 Genus Dracorex Q9 1 Dracorex hogwartsia Q10 1 dragon king of Hogwarts S6Q1 1 Species neanderthalensis Q2 1 Eurasia Q3 1 Any of the following: neanderthals are stockier, had shorter legs, had bigger bodies Q4 1 Genus Dimetrodon Q5 1 Within 50 Ma: 295-272 Ma Q6 1 Sphenacodontidae Q7 1 Genus Smilodon Q8 1 Class Mammalia Q9 1 Pleistocene epoch Q10 1 1842 S7Q1 1 Shale Q2 1 Compaction Q3 1 Clastic Q4 1 Sandstone Q5 1 Any of the following: river, ocean, glacier, desert deposits Q6 1 clastic Q7 1 Coquina Q8 1 FALSE Q9 1 Any of the following: shells of molluscs, trilobites, brachiopods, or other invertebrates Q10 1 organic. -

Geological Magazine
THE GEOLOGICAL MAGAZINE. No. LIU—NOVEMBER, 1868. ABTICLES. I.—ON A NEWLY-DISCOVERED LONG-EYED CALTJIMNE FEOM THE WENLOOK LIMESTONE, DUDLEY. BY HBNKT •WOODWABD, F.G.S., F.Z.S., of the British Museum. [PLATE XXI.] T is more than a century since the " Dudley Locust," or ' Trilo- I bite,' was first figured and described,1 and the locality where it it is found, is rendered famous by the researches and writings of Sedgwick, Phillips, Forbes, Murchison, Salter, Davidson, and a host of other geologists and palaeontologists, who have been attracted thither at various times by the graiad' geological features of the dis- trict or by the matchless beauty and endless diversity of its well- preserved organic remains. • • • * - Nor have the advantages, which this locality offers, been lost sight of by the members of the Dudley and Midland Geological Society, whose well-stored cabinets attest the earnest interest they all take in procuring and preserving the choicest Corals, Mollusca, Crinoids, and Trilobites, which the Wenlock Limestone and Shale so abund- antly afford. To one of these gentlemen, Mr. E. Hollier, of Dudley, I am in- debted for the opportunity of examining and describing the remark- able Trilobite which forms the subject of this communication. From the time of the establishment of the genus Calymene by Brongniart in 1822,* the " Dudley Trilobite " may be said to have been very well known, described, and figured, and its portrait has appeared in almost every geological work in which fossils have been noticed from that time down to the present day. For the best description and illustration of this and many other genera of British Trilobites we are indebted to Mr. -
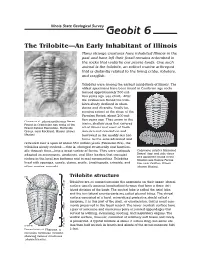
Geobit 6: the Trilobite
Illinois State Geological Survey Geobit 6 The Trilobite—An Early Inhabitant of Illinois Many strange creatures have inhabited Illinois in the past and have left their fossil remains entombed in the rocks that underlie our prairie lands. One such animal is the trilobite, an extinct marine arthropod that is distantly related to the living crabs, lobsters, and crayfish. Trilobites were among the earliest inhabitants of Illinois. The oldest specimens have been found in Cambrian age rocks formed approximately 500 mil- lion years ago (see chart). After the Ordovician Period the trilo- bites slowly declined in abun- dance and diversity, finally be- coming extinct at the close of the Permian Period, about 200 mil- lion years ago. They swam in the Ceraurus cf. pleurexanthemus Green. Found in Ordovician age rocks of the warm, shallow seas that covered Grand Detour Formation, Platteville all of Illinois and most of North Group, near Rockford, Illinois (shown America and crawled on and lifesize). burrowed in the muddy sea bot- toms. As the seas advanced and retreated over a span of about 350 million years (Paleozoic Era), the trilobites slowly evolved —that is, changed structurally and function- ally through time—into a great variety of forms. They were variously Calymene celebra Raymond. adapted as scavengers, predators, and filter feeders that occupied Dorsal (top) and side views of a specimen found in the niches in the level sea bottoms and in reef communities. Trilobites Silurian age Racine Forma- lived with sponges, corals, clams, snails, brachiopods, crinoids, and tion near Grafton, Illinois other marine animals. (shown lifesize). ��� Trilobite structure � � � Trilobites are so named because the segments on their upper (dorsal) � � � surface usually possess longitudinal furrows that form a three (tri-) � � lobed division of the body. -

From-1\ the Mi41ddle -D.Evoni)*;An O)F No(Rthnamerica
S!YSTiEM\ATICS ANID EVO(LUJT(ION* O)F PH-ACOPS RAN (GEEN, 1832) AN1*D PHIACOPS IOWVEN)SIS DELO, 1935 (TR.ILoBITA) FROM-1\ THE MI41DDLE -D.EVONI)*;AN O)F NO(RTHNAMERICA NILES ELDREDGE BULLETIN OF THE AMERICAN MUSEUM OF NATURAL HISTORY VOLUME 147:ARTICLE 2 NEW YORK:1,972 SYSTEMATICS AND EVOLUTION OF PHACOPS RANA (GREEN, 1832) AND PHA COPS IOWENSIS DELO, 1935 (TRILOBITA) FROM THE MIDDLE DEVONIAN OF NORTH AMERICA NILES ELDREDGE Assistant Curator, Department ofInvertebrate Paleontology The American Museum ofNatural History BULLETIN OF THE AMERICAN MUSEUM OF NATURAL HISTORY VOLUME 147: ARTICLE 2 NEW YORK : 1972 BULLETIN OF THE AMERICAN MUSEUM OF NATURAL HISTORY Volume 147, article 2, pages 45-114, figures 1-28, tables 1-9 Issued February 14, 1972 Price: $2.70 a copy Printed in Great Britain by Lund Humphries CONTENTS ABSTRACT....................................49 INTRODUCTION..................................51 Abbreviations.................................52 MIDDLE DEVONIAN STRATIGRAPHY..........................5 MORPHOLOGY AND RELATIONSHIPS OF THE BiOSPECIES Plhacops rana (Green, 1832) and Phacops iowensisDelo, 1935...............................56 Systematic Paleontology.............................58 PhacopsiowensisDelo, 1935..........................58 Phacopsrana (Green, 1832)...........................59 The Affinities ofPhacops rana and Phacops iowensis...................59 Previous Work on Middle Devonian Phacopid Taxa in North America..........60 ANALYTICAL TECHNIQUES ...............61 ONTOGENY OF THE CEPHALON OF Phacops rana.....................65 -

Digestive Structures in Ordovician Trilobites Colpocoryphe and Flexicalymene from the Barrandian Area of Czech Republic
Estonian Journal of Earth Sciences, 2015, 64, 4, 255–266 doi: 10.3176/earth.2015.32 Digestive structures in Ordovician trilobites Colpocoryphe and Flexicalymene from the Barrandian area of Czech Republic Oldřich Fatkaa, Petr Budilb and Martin Davidc a Charles University, Faculty of Sciences Institute of Geology and Palaeontology, Albertov 6, 128 43, Prague 2, Czech Republic; [email protected] b Czech Geological Survey, Klárov 3, 118 21 Prague 1, Czech Republic; [email protected] c Rožmberská 613/10, Praha 9-Kyje, 198 00, Czech Republic; [email protected] Received 21 July 2015, accepted 18 September 2015 Abstract. Two recently discovered specimens of the abundant calymenoid trilobite Colpocoryphe Novák in Perner, 1918 from the Middle Ordovician Šárka Formation and one specimen of Flexicalymene (Flexicalymene) pragensis Vaněk & Vokáč, 1997 from the Upper Ordovician Bohdalec Formation, all from the Prague Basin, display remains of the digestive system. In Colpocoryphe, an internal mould of an articulated exoskeleton contains a post-stomach part of the alimentary canal preserved through the narrow axial region of the occipital ring, all thoracic segments as well as in the axial part of the pygidial shield. The anterior part of the digestive system is poorly known as the specimen shows the hypostome preserved in situ and the space between the glabella and the hypostome is represented by an empty cavity associated with probable remains of gut diverticulae on both sides of the cephalon. The second, incomplete specimen consists of five posterior thoracic segments articulated with the pygidium. The axial region of this specimen is preserved with a clearly discernible segmented intestine which terminates at the axial tip and then bends ventrally.