A Hierarchical Analysis of the Genetic Structure of an Aquatic Insect 2 Bungona
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
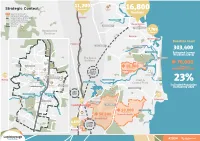
Strategic Context
11,200 Residents The Airport Strategic Context Project 16,800 Residents Major Development Area Recreational/Tourism Zones Nambour Landsborough Local Plan Area Boundary MAROOCHY RIVER Landsborough Study Area Area of Significant Population Maroochydore Water Body MAROOCHYDORE RD Train Line SUNSHINE MOTORWAY Kondalilla 7,700 National Park Hinterland & Elevation Residents Buderim Mooloolaba Sunshine Coast Palmwoods BRUCE HIGHWAY LAKE BAROON SUNSHINE MOTORWAY 303,400 Dularcha MOOLOOLAH RIVER National Park Estimated Current University of the Residents (2016) Sunshine Coast NICKLIN WAY The Space Mooloolah River Between National Park 70,000 MALENY STREET LAKE KAWANA Landsborough 18,000 Estimated Sportsgrounds Estimated Residents Residents by 2026 40MIN Palmview BY CAR TO Landsborough State FUTURE CITY Primary School Peace Memorial CENTRE Sunshine Coast 3,700 CRIBB STREET University Hospital Residents Park CALOUNDRA STREET Maleny Landsborough Train Station Pioneer Park MOOLOOLAH RIVER Coast & Landsborough Bus Station EWEN MADDOCK DAM 23% Coastal Plain Landsborough STEVE IRWIN WAY Estimated Population Landsborough Police Station CALOUNDRA ROAD Increase by 2026 Beerburrum15MIN Beerwah State Forest LITTLE ROCKY CREEK StateBY Forest CAR TO MELLUM CREEK MALENY 3,900 Residents Big Kart Track 3,800 Caloundra STEVE IRWIN WAY Rocky Creek Residents STEVE IRWIN WAY Camp Site Landsborough BRUCE HIGHWAY Skippy Park 50,000 LAKE MAGELLAN Estimated Residents Caloundra South Australia Zoo 50,000 Estimated Residents Beerwah East 6,800 1.25HRS Residents BY CAR TO BRISBANE Beerwah Setting the The rural township of Scene Landsborough is situated at the southern entrance of the Blackall Range with the areas surrounding the township being rural residential and rural lands. This regional inter-urban break is a significant feature that frames the township and shapes its identity. -

April 8, 2020 | Edition 200 |
Sunshine Valley GazetteFREE Local Matters | 10,000 Copies Fortnightly | Wednesday April 8, 2020 | Edition 200 | www.sunshinevalleygazette.com.au Distributed to Palmwoods, Woombye, Nambour, Chevallum, Mooloolah, Editorial: [email protected] 0409 424 487 Eudlo, Hunchy, Yandina, Montville, Flaxton, Mapleton, Maleny. Advertising: [email protected] 0400 069 711 “We have changed our offerings to comply with restrictions & support our customers & suppliers.” The little cafe with a big heart ... Sweethearts Cafe owner Sharon Knight and her son Adam (holding new fruit and veggie box) & daughter Beth with baby Rex ... “Being a family business we just have to survive and we have a huge passion for this small community and surrounds.” Getting on with business AS businesses across the hinterland adapt just so very grateful for the support from we have such a strong belief in the power of “We now supply weekly fruit and veggie to cope with the impacts of COVID-19 one our beautiful customers who are certainly community. That is certainly being put to the boxes, sourced locally, and from this week small cafe in the tiny town of Eudlo stands a loyal bunch. Anything that we can do to test right now, and we have never been more we will be adding other fresh staples such as as a beacon of inspiration. make life easier for them we will as it helps determined to make this work. beautiful local bacon and small goods, meat Since opening three years ago, the aptly us keep our doors open.” “We take the COVID-19 restrictions very packs, Sunshine Coast milk, and juice. -

Tom Petrie's Reminiscences
I TOM PETRIE'S REMINISCENCES OF EARLY QUEENSLAND (Dating from 1837.) RECORDED BY HIS DAUGHTER. BRISBANE: WATSON , FERGUSON & CO.. 1904. [COPYRIGHT.] This is a blank page To MY FATHER, TOM PETRIE, WHOSE FAITHFUL MEMORY HAS SUPPLIED THE MATERIAL FOR THIS BOOK. PRINTED BY WATSON, FERGUSON &' CO. QUEEN ST., BRISBANE. This is a blank page This is a blank page NOTE. THE greater portion of the contents of this book first ap- peard in the " Queenslander " in the form of articles, and when those referring to the aborigines were pubished, Dr. Roth, author of " Ethnological Studies," etc., wrote the following letter to that paper :- TOM PETRIE' S REMINISCENCES (By C.C.P.) TO THE EDITOR. SIR,-lt is with extreme interest that I have perused the remarkable series of articles appearing in the Queenslander under the above heading, and sincerely trust that they will he subsequently reprinted. The aborigines of Australia are fast dying out, and with them one of the most interesting phases in the history and development of man. Articles such as these, referring to the old Brisbane blacks, of whom I believe but one old warrior still remains, are well worth permanently recording in convenient book form-they are, all of them, clear, straight-forward statements of facts- many of which by analogy, and from early records, I have been able to confirm and verify-they show an intimate and profound knowledge of the aboriginals with whom they deal, and if only to show with what diligence they have been written, the native names are correctly, i.e., rationally spelt. -

SUNSTATE CHARTERS Day Tour & Speciality Tour Brochure 2017
SUNSTATE CHARTERS Day Tour & Speciality Tour Brochure 2017 PO Box1232 Eagle Farm QLD 4009. Tel: 07 3260 1666 Fax: 07 3260 1966 [email protected] www.sunstatecoaches.com.au SUNSTATE CHARTERS PTY LTD :ABN 45 010 609 757 Sunstate Charters Tours for 2017 Hello travellers! Sunstate Charters present’s our Day & Speciality Touring brochure for 2017. To freshen up the touring we have been hard at work creating new tours and updating old favourites to increase the choice to you, our valued clients. This year’s big change is the introduction of the speciality touring range, “Sunstate Speciality Tours” listed in the back of the brochure. The speciality touring range includes Coach Travel, Sightseeing, Accommodation and listed meals. Travelling with an experienced Coach Captain who will provide you with a unique travelling experience by providing informative commentary along the way. As always, Sunstate endeavour to provide an enjoyable and safe day out. So to help make your day a special one, please do not hesitate to inform us of any special requirements. Remember we offer a “Create your own adventure” option. This offers the group total flexibility on your own designed and organised itinerary. Present your itinerary to Sunstate Charters and we will quote a price that includes the coach, driver and morning tea (optional) only. This option can mean a cheaper day out for the group depending on your choice of destinations, lunch and tour options. Now where would you like Sunstate Charters to take you today? Discover Redcliffe The day starts with a leisurely drive to the Clontarf Visitors Centre, where we pick up our local guide for the day, before heading over to Pelican Park for morning tea overlooking Bramble Bay. -

Blackall Range Range Blackall
top left); © Robert Ashdown (top right) (top Ashdown Robert © left); top Adam Creed © Qld Govt Qld © Creed Adam © Ross Naumann, QPWS Volunteer (above and and (above Volunteer QPWS Naumann, Ross © while you’re walking. you’re while 1km with numerous switchbacks to descend the 100m into into 100m the descend to switchbacks numerous with 1km Steven Nowakowski © Toursim and Events Qld Events and Toursim © Nowakowski Steven Coast Hinterland Great Walk Great Hinterland Coast barred frog (top right); lace monitor (above) monitor lace right); (top frog barred Gheerulla Bluff, Sunshine Sunshine Bluff, Gheerulla View over Mapleton over View the wompoo fruit-doves’ ‘wallock-a-woo’ and ‘book-a-roo’, ‘book-a-roo’, and ‘wallock-a-woo’ fruit-doves’ wompoo the From the rock pools, the Kondalilla Falls circuit winds over over winds circuit Falls Kondalilla the pools, rock the From , great great , fasciolatus Mixophyes left); (top Figbird Steep climb up the the up climb Steep that carry through the valley. Be sure to listen for a call, like like call, a for listen to sure Be valley. the through carry that Kondalilla Falls circuit Falls Kondalilla 2 the on stairs 300 over are There . catch a glimpse, then you will certainly enjoy their birdsongs birdsongs their enjoy certainly will you then glimpse, a catch lush rainforest to the base of the falls. falls. the of base the to rainforest lush binoculars to get a close-up look at the birdlife. If you can’t can’t you If birdlife. the at look close-up a get to binoculars cool, through down on and grasstrees, and forest eucalypt forest is an easy walk for families with children. -

Sunshine Coast | Queensland Historical Atlas
Search Home Quintessential Queensland Distance Division Dreaming Development Home » Development » Transformation Sunshine Coast By: Chris McConville The Sunshine Coast is one of Australia’s most rapidly changing regions. On the one hand inter-state migration is transforming once distinctive coastal and farming townships into a uniform suburban expanse. On the other, climatic transitions are reshaping the wide beaches and pristine waterways. As a result the once clear northern boundary, formed in the broad sweep of the Noosa river estuary, is now fudged, as tourist development inserts itself on the forested north bank of the river. The Sunshine Coast, c1984 southern borderland of the Sunshine Coast meanders between the marine park of Pumicestone Passage, commercial pine plantations and the conical peaks and fruit farms of the Glasshouse Mountains. These form a precarious divide between the distinctiveness of the Sunshine Coast as a Suburban portions of Noosa and Tewantin, 1944 region and a fast approaching northern boundary of suburban Brisbane. In the main a sandy lowland, the Sunshine Coast runs from north of Brisbane to the Noosa River. It is bounded to the east by surf beaches and the Coral Sea, to the west the abrupt escarpment of the Blackall Range Sunshine Coast, 2000 defines the inland margins of the Coast. Here dense forest, few made roads and sparsely populated green valleys demarcate a natural order quite distinct from the breathless rebuilding of the coast. The recently-created Sunshine Coast Regional Council runs across three estuarine environments of the Mooloolah, Maroochy and Noosa Rivers. These coastal flat lands often swampy and in part below sea level were formed over thousands of years by sand washed ashore along coastal currents. -

The Blackall Ra Glass House Mountains
rt 4 Aussie World To Palmwoods C Mary Valley a Information Centres Legend d Rattler n Ettamogah Pub urg Road e 3 Brandenb r Montville Information Centre GLASS HOUSE B THE Winery Moo Phone: 07 5478 5544 d loo Mooloolah a la Mooloolah o h Toilets R C Maleny Visitor Information Centre TRAIN STATION on River National Park Jowarra Section e n ect Phone: 07 5499 9033 The Ginger Factory g ion To Caloundra d Ro i ad Lookout Ph: 1800 067 686 MOUNTAINS y BLACKALL RANGE R a Kenilworth Information Centre l e w n Ewen Maddock Dam h n Phone: 07 5446 0122 g u y Picnic Area a i T in W Steve Irw H Dularcha e To Noosa Mapleton Community Information Sunshine Coast Hinterland c Sunshine Coast National u Police Station Palmwoods r & Eumundi Centre Phone: 07 5478 6381 Park B To Maleny, Montville, the walks Glass House Mountains Information S.C. Hinterland Maleny Visitor and villages of The Blackall Range Hinterland Great Walk LEGEND Centre Phone: 07 5438 7220 Big Kart Track Info Centre (separate map available) The Geology & European History Toilets www.hinterlandtourism.com.au Landsborough 07 5499 9033 La The Glass House Mountains are subterranean intrusions of Museum nd Lookout s d magna formed throughout the area some 25 million years ago. b a o ) o r e To Imbil To Eumundi To Yandina d o Overtime the softer rock surrounding these intrusions has been v R i u a Picnic Area r b gh - Ma o o l D r n e eroded away. -

Mapleton Has Extensive Areas of Red Basalt Soil, the Result of Volcanic Activity South West Mapleton of Maleny Between 31 and 27 Million Years Ago
Great Walk, National Parks, Walks, Waterfalls Mapleton National Park, formerly Mapleton and Gheerulla Falls is on the Great Walk and can www.npsr.qld.gov.au Cooloolabin (Timber) Reserves, covers 10,426ha also be accessed from Delicia Road. Gheerulla is and was gazetted as a National Park on World a contraction of the words kirar nulla of the Waka Sunshine Coast Hinterland Great Walk Environment Day 5 June 2011. It contains about half people and means ‘dry or empty creek’. Towering bunyas, rushing waterfalls and rugged, of the Great Walk. Several shorter walks including scarred landscapes are some of the natural wonders the Turpentine Trail (7.8km return), Piccabeen Circuit to be discovered on this 58km walk. There are (6.7km) and Kureelpa Falls (8.5km) start from the several walks from two to seven hours. The Great Day Use Area. A short Pilularis Forest Walk (400m Walk map can be purchased from the Mapleton return) starts from Leafy Lane trailhead. The Linda Information Centre. Garrett Circuit (700m return) and Gheerulla Falls (2.5km return) are accessed from Delicia Road. Mapleton Falls National Park, covering 26ha, GH became a recreation reserve in 1893 and a National There are 85km of horse riding trails,a 26km trail Park in 1975. The 1.3km Wompoo Circuit walk starts bike track and two backpackers campsites, Ubajee Baxter Creek Falls is on a short sidewalk from the from the lookout and winds through eucalypts and and Thilba Thalba. The Day Use Area has picnic Great Walk near a metal suspension bridge crossing rainforest. Peregrine falcon may be sighted. -

Revised List of Queensland Birds
Records of the Western Australian Museum Supplement 0 19. 1984 Revised List ofQueensland Birds G.M.Storr ,~ , , ' > " Records of the Western Australian Museum Supplement No. 19 I $ I,, 1 > Revised List oflQueensland Birds G. M. Storr ,: i, Perth 1984 'j t ,~. i, .', World List Abbreviation: . Rec. West. Aust. Mus. Suppl. no. 19 Cover Palm Cockatoo (Probosciger aterrimus), drawn by Jill Hollis. © Western Australian Museum 1984 I ISBN 0 7244 8765 4 Printed and Published by the Western Australian Museum, j Francis Street, Perth 6000, Western Australia. TABLE OF CONTENTS Page Introduction. ...................................... 5 List of birds. ...................................... 7 Gazetteer ....................................... .. 179 3 INTRODUCTION In 1967 I began to search the literature for information on Queensland birds their distribution, ecological status, relative abundance, habitat preferences, breeding season, movements and taxonomy. In addition much unpublished information was received from Mrs H.B. Gill, Messrs J.R. Ford, S.A. Parker, R.L. Pink, R.K. Carruthers, L. Neilsen, D. Howe, C.A.C. Cameron, Bro. Matthew Heron, Dr D.L. Serventy and the late W.E. Alexander. These data formed the basis of the List of Queensland birds (Stort 1973, Spec. Pubis West. Aust. Mus. No. 5). During the last decade the increase in our knowledge of Queensland birds has been such as to warrant a re-writing of the List. Much of this progress has been due to three things: (1) survey work by J.R. Ford, A. Gieensmith and N.C.H. Reid in central Queensland and southern Cape York Peninsula (Ford et al. 1981, Sunbird 11: 58-70), (2) research into the higher categories ofclassification, especially C.G. -

Conondale Range Conservation Issue One Newsletter April 2019
Conondale Range Conservation PO Box 150 KENILWORTH 4574 Newsletter April 2019 Issue one Yabba National Park The Yabba Falls Scenic Reserve was gazetted in the 1920s, predating all national parks in the Conondales region. A year or two back, Imbil-based geographer Ian Stehbens made a very detailed submission for a national ark around Yabba Falls, Kingaham Falls and the gorges associated with each. Ian sought support from NPAQ who were impressed not just with the country involved but with the background work he had put in. He sought wider support, from the Royal Geographical Society who spent a weekend exploring there last August, and he sought support from us, Conondale Range Conservation, from Hinterland Bush Links and from SCEC, the Sunshine Coast Environment Council. Looking at other state forests surrounding Ian’s proposal it was plain that many would have been on the verge of transitioning into national parks had Campbell Newman not axed the SEQFA. Both Conondale NP to the south and Wrattens NP to the north had several small outliers already converted to NP under the SEQFA. And given that the government had a commitment to a substantial increase in national parks, it was plain that the area around Yabba Falls provided an enormous opportunity. NPAQ had Tyler Hood research and write up a lengthier proposal and conducted a number of flora and fauna surveys of the area in conjunction with Birds Queensland and others. As well as providing connectivity between NP outliers, the proposed Yabba NP would provide a north-south link between the bulk of Wrattens NP to the north and that of Conondale NP to the south. -

Thematic History of the Sunshine Coast Sunshine Coast Heritage Study Sunshine Coast Council August 2020
Thematic History of the Sunshine Coast Sunshine Coast Heritage Study Sunshine Coast Council August 2020 Converge Heritage + Community Contact details are: Simon Gall Converge Heritage + Community ABN:71 366 535 889 GPO Box 1700, Brisbane, 4001 Tel: (07) 3211 9522 Email: [email protected] Copyright © 2018 Document Verification Project Sunshine Coast Heritage Study Project Number 16031C Document Title Thematic History of the Sunshine Coast FINAL_14-02-2018 File Location Share Point Client Sunshine Coast Council Version history Revision Date Nature of revision Prepared by Authorised by 0 15/02/2016 Draft report BR, CB CB 1 14/02/2018 Final report CB - 2 Thematic History of the Sunshine Coast | 2 Contents 1 Introduction ........................................................................................................................ 7 1.1 Project Background ....................................................................................................................... 7 1.2 Purpose of this report ................................................................................................................... 7 1.3 Methodology ................................................................................................................................. 8 1.4 Sources .......................................................................................................................................... 9 1.5 Aboriginal cultural heritage and scope of document .....................Error! Bookmark not defined. 1.6 Authorship -

English Cop16 Prop. 40 CONVENTION on INTERNATIONAL TRADE in ENDANGERED SPECIES OF
Original language: English CoP16 Prop. 40 CONVENTION ON INTERNATIONAL TRADE IN ENDANGERED SPECIES OF WILD FAUNA AND FLORA ____________________ Sixteenth meeting of the Conference of the Parties Bangkok (Thailand), 3-15 March 2013 CONSIDERATION OF PROPOSALS FOR AMENDMENT OF APPENDICES I AND II A. Proposal Delist the extinct Rheobatrachus silus from Appendix II in accordance with the Resolution Conf. 9.24 (Rev. CoP 15). The species does not meet the trade criteria (Annexes 2a and 2b) for inclusion in Appendix II. B. Proponent Australia*, as requested by the Animals Committee, to delete the species from Appendix II (AC 26 WG1 Doc. 2). C. Supporting statement 1. Taxonomy 1.1 Class: Amphibia 1.2 Order: Anura 1.3 Family: Myobatrachidae 1.4 Species: Rheobatrachus silus Liem, 1973 1.5 Scientific synonyms: none 1.6 Common names: English: southern gastric-brooding frog, southern platypus frog Dutch zuidelijke maagbroedkikker French: grenouille plate à incubation gastrique German Australische magenbrüterfrosch Spanish: rana incubadora gástrica de Australia Swedish magruvargroda 1.7 Code numbers: 2. Overview At the 24th meeting of the Animals Committee (Geneva, April 2009), the southern gastric-brooding frog (Rheobatrachus silus) was selected for the periodic review of animal species included in the CITES Appendices. At their 26th meeting (Geneva, March 2012), the Animals Committee recommended that the southern gastric-brooding frog be removed from Appendix II (AC 26 WG1 Doc. 2). The recommendation was made based on information provided by the Australian CITES Scientific Authority. * The geographical designations employed in this document do not imply the expression of any opinion whatsoever on the part of the CITES Secretariat or the United Nations Environment Programme concerning the legal status of any country, territory, or area, or concerning the delimitation of its frontiers or boundaries.