The Carpus and Tarsus of the Early Permian Synapsid Sphenacodon Ferox (Eupelycosauria: Sphenacodontidae)
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Distributions of Extinction Times from Fossil Ages and Tree Topologies: the Example of Some Mid-Permian Synapsid Extinctions Gilles Didier, Michel Laurin
Distributions of extinction times from fossil ages and tree topologies: the example of some mid-Permian synapsid extinctions Gilles Didier, Michel Laurin To cite this version: Gilles Didier, Michel Laurin. Distributions of extinction times from fossil ages and tree topologies: the example of some mid-Permian synapsid extinctions. 2021. hal-03258099v2 HAL Id: hal-03258099 https://hal.archives-ouvertes.fr/hal-03258099v2 Preprint submitted on 20 Sep 2021 HAL is a multi-disciplinary open access L’archive ouverte pluridisciplinaire HAL, est archive for the deposit and dissemination of sci- destinée au dépôt et à la diffusion de documents entific research documents, whether they are pub- scientifiques de niveau recherche, publiés ou non, lished or not. The documents may come from émanant des établissements d’enseignement et de teaching and research institutions in France or recherche français ou étrangers, des laboratoires abroad, or from public or private research centers. publics ou privés. Distributions of extinction times from fossil ages and tree topologies: the example of some mid-Permian synapsid extinctions Gilles Didier1 and Michel Laurin2 1 IMAG, Univ Montpellier, CNRS, Montpellier, France 2 CR2P (\Centre de Pal´eontologie { Paris"; UMR 7207), CNRS/MNHN/SU, Mus´eumNational d'Histoire Naturelle, Paris, France September 16, 2021 Abstract Given a phylogenetic tree that includes only extinct, or a mix of extinct and extant taxa, where at least some fossil data are available, we present a method to compute the distribution of the extinction time of a given set of taxa under the Fossilized-Birth-Death model. Our approach differs from the previous ones in that it takes into account (i) the possibility that the taxa or the clade considered may diversify before going extinct and (ii) the whole phylogenetic tree to estimate extinction times, whilst previous methods do not consider the diversification process and deal with each branch independently. -
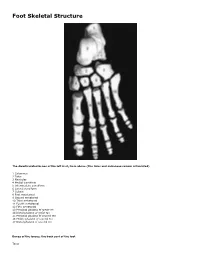
Skeletal Foot Structure
Foot Skeletal Structure The disarticulated bones of the left foot, from above (The talus and calcaneus remain articulated) 1 Calcaneus 2 Talus 3 Navicular 4 Medial cuneiform 5 Intermediate cuneiform 6 Lateral cuneiform 7 Cuboid 8 First metatarsal 9 Second metatarsal 10 Third metatarsal 11 Fourth metatarsal 12 Fifth metatarsal 13 Proximal phalanx of great toe 14 Distal phalanx of great toe 15 Proximal phalanx of second toe 16 Middle phalanx of second toe 17 Distal phalanx of second toe Bones of the tarsus, the back part of the foot Talus Calcaneus Navicular bone Cuboid bone Medial, intermediate and lateral cuneiform bones Bones of the metatarsus, the forepart of the foot First to fifth metatarsal bones (numbered from the medial side) Bones of the toes or digits Phalanges -- a proximal and a distal phalanx for the great toe; proximal, middle and distal phalanges for the second to fifth toes Sesamoid bones Two always present in the tendons of flexor hallucis brevis Origin and meaning of some terms associated with the foot Tibia: Latin for a flute or pipe; the shin bone has a fanciful resemblance to this wind instrument. Fibula: Latin for a pin or skewer; the long thin bone of the leg. Adjective fibular or peroneal, which is from the Greek for pin. Tarsus: Greek for a wicker frame; the basic framework for the back of the foot. Metatarsus: Greek for beyond the tarsus; the forepart of the foot. Talus (astragalus): Latin (Greek) for one of a set of dice; viewed from above the main part of the talus has a rather square appearance. -
Reptile Family Tree
Reptile Family Tree - Peters 2015 Distribution of Scales, Scutes, Hair and Feathers Fish scales 100 Ichthyostega Eldeceeon 1990.7.1 Pederpes 91 Eldeceeon holotype Gephyrostegus watsoni Eryops 67 Solenodonsaurus 87 Proterogyrinus 85 100 Chroniosaurus Eoherpeton 94 72 Chroniosaurus PIN3585/124 98 Seymouria Chroniosuchus Kotlassia 58 94 Westlothiana Casineria Utegenia 84 Brouffia 95 78 Amphibamus 71 93 77 Coelostegus Cacops Paleothyris Adelospondylus 91 78 82 99 Hylonomus 100 Brachydectes Protorothyris MCZ1532 Eocaecilia 95 91 Protorothyris CM 8617 77 95 Doleserpeton 98 Gerobatrachus Protorothyris MCZ 2149 Rana 86 52 Microbrachis 92 Elliotsmithia Pantylus 93 Apsisaurus 83 92 Anthracodromeus 84 85 Aerosaurus 95 85 Utaherpeton 82 Varanodon 95 Tuditanus 91 98 61 90 Eoserpeton Varanops Diplocaulus Varanosaurus FMNH PR 1760 88 100 Sauropleura Varanosaurus BSPHM 1901 XV20 78 Ptyonius 98 89 Archaeothyris Scincosaurus 77 84 Ophiacodon 95 Micraroter 79 98 Batropetes Rhynchonkos Cutleria 59 Nikkasaurus 95 54 Biarmosuchus Silvanerpeton 72 Titanophoneus Gephyrostegeus bohemicus 96 Procynosuchus 68 100 Megazostrodon Mammal 88 Homo sapiens 100 66 Stenocybus hair 91 94 IVPP V18117 69 Galechirus 69 97 62 Suminia Niaftasuchus 65 Microurania 98 Urumqia 91 Bruktererpeton 65 IVPP V 18120 85 Venjukovia 98 100 Thuringothyris MNG 7729 Thuringothyris MNG 10183 100 Eodicynodon Dicynodon 91 Cephalerpeton 54 Reiszorhinus Haptodus 62 Concordia KUVP 8702a 95 59 Ianthasaurus 87 87 Concordia KUVP 96/95 85 Edaphosaurus Romeria primus 87 Glaucosaurus Romeria texana Secodontosaurus -

Origin and Beyond
EVOLUTION ORIGIN ANDBEYOND Gould, who alerted him to the fact the Galapagos finches ORIGIN AND BEYOND were distinct but closely related species. Darwin investigated ALFRED RUSSEL WALLACE (1823–1913) the breeding and artificial selection of domesticated animals, and learned about species, time, and the fossil record from despite the inspiration and wealth of data he had gathered during his years aboard the Alfred Russel Wallace was a school teacher and naturalist who gave up teaching the anatomist Richard Owen, who had worked on many of to earn his living as a professional collector of exotic plants and animals from beagle, darwin took many years to formulate his theory and ready it for publication – Darwin’s vertebrate specimens and, in 1842, had “invented” the tropics. He collected extensively in South America, and from 1854 in the so long, in fact, that he was almost beaten to publication. nevertheless, when it dinosaurs as a separate category of reptiles. islands of the Malay archipelago. From these experiences, Wallace realized By 1842, Darwin’s evolutionary ideas were sufficiently emerged, darwin’s work had a profound effect. that species exist in variant advanced for him to produce a 35-page sketch and, by forms and that changes in 1844, a 250-page synthesis, a copy of which he sent in 1847 the environment could lead During a long life, Charles After his five-year round the world voyage, Darwin arrived Darwin saw himself largely as a geologist, and published to the botanist, Joseph Dalton Hooker. This trusted friend to the loss of any ill-adapted Darwin wrote numerous back at the family home in Shrewsbury on 5 October 1836. -

2018 NMGS Spring Meeting: Abstract-748
FIRST DISCOVERY OF A TETRAPOD BODY FOSSIL IN THE LOWER PERMIAN YESO GROUP, CENTRAL NEW MEXICO Emily D. Thorpe1, Spencer G. Lucas2, David S. Berman3, Larry F. Rinehart2, Vincent Santucci4 and Amy C. Henrici3 1 POBox 147, Morrisonville, WI, 53571, [email protected] 2New Mexico Museum of Natural History and Science, 1801 Mountain Road N. W., Albuquerque, NM, 87104 3Carnegie Museum of Natural History, 4400 Forbes Ave, Pittsburgh, PA, 15213 4National Parks Service, 1849 C Street, NW, Washington, DC, 20240, United States The lower Permian Yeso Group records arid coastal plain, shallow marine, and evaporitic deposition across much of central New Mexico. Generally considered to have few fossils, recent study of Yeso Group strata has discovered a diverse fossil record of marine micro-organisms (mostly algae and foraminiferans), terrestrial plants, and tetrapod footprints. We report here the first discovery of a tetrapod body fossil in the Yeso Group—a partial skeleton of a basal synapsid, varanopidae eupelycosaur. The fossil is the natural casts of bones in two pieces, part and counterpart, that were preserved in a sandstone bed of the lower part of the Arroyo de Alamillo Formation in the southern Manzano Mountains. The fossil-bearing sandstone is fine-grained, quartz rich, and pale reddish brown to grayish red unweathered, weathers to blackish red, and is in part encrusted by white caliche. The casts preserve part of the pelvis(?), 18 caudal vertebral centra, both femora and tibia-fibulae, and most of the pedes, largely in close articulation, of a single individual. The skeleton is of a relatively small (femur length = 62 mm, total length of the preserved cast from the pelvis to tip of the incomplete tail = 325 mm) and gracile eupelycosaur most similar to Varanops. -

Morphology, Phylogeny, and Evolution of Diadectidae (Cotylosauria: Diadectomorpha)
Morphology, Phylogeny, and Evolution of Diadectidae (Cotylosauria: Diadectomorpha) by Richard Kissel A thesis submitted in conformity with the requirements for the degree of doctor of philosophy Graduate Department of Ecology & Evolutionary Biology University of Toronto © Copyright by Richard Kissel 2010 Morphology, Phylogeny, and Evolution of Diadectidae (Cotylosauria: Diadectomorpha) Richard Kissel Doctor of Philosophy Graduate Department of Ecology & Evolutionary Biology University of Toronto 2010 Abstract Based on dental, cranial, and postcranial anatomy, members of the Permo-Carboniferous clade Diadectidae are generally regarded as the earliest tetrapods capable of processing high-fiber plant material; presented here is a review of diadectid morphology, phylogeny, taxonomy, and paleozoogeography. Phylogenetic analyses support the monophyly of Diadectidae within Diadectomorpha, the sister-group to Amniota, with Limnoscelis as the sister-taxon to Tseajaia + Diadectidae. Analysis of diadectid interrelationships of all known taxa for which adequate specimens and information are known—the first of its kind conducted—positions Ambedus pusillus as the sister-taxon to all other forms, with Diadectes sanmiguelensis, Orobates pabsti, Desmatodon hesperis, Diadectes absitus, and (Diadectes sideropelicus + Diadectes tenuitectes + Diasparactus zenos) representing progressively more derived taxa in a series of nested clades. In light of these results, it is recommended herein that the species Diadectes sanmiguelensis be referred to the new genus -

Distributions of Extinction Times from Fossil Ages and Tree Topologies: the Example of Some Mid-Permian Synapsid Extinctions
bioRxiv preprint doi: https://doi.org/10.1101/2021.06.11.448028; this version posted June 11, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder. All rights reserved. No reuse allowed without permission. Distributions of extinction times from fossil ages and tree topologies: the example of some mid-Permian synapsid extinctions Gilles Didier1 and Michel Laurin2 1IMAG, Univ Montpellier, CNRS, Montpellier, France 2CR2P (“Centre de Recherches sur la Paléobiodiversité et les Paléoenvironnements”; UMR 7207), CNRS/MNHN/UPMC, Sorbonne Université, Muséum National d’Histoire Naturelle, Paris, France June 11, 2021 Abstract Given a phylogenetic tree of extinct and extant taxa with fossils where the only temporal infor- mation stands in the fossil ages, we devise a method to compute the distribution of the extinction time of a given set of taxa under the Fossilized-Birth-Death model. Our approach differs from the previous ones in that it takes into account the possibility that the taxa or the clade considered may diversify before going extinct, whilst previous methods just rely on the fossil recovery rate to estimate confidence intervals. We assess and compare our new approach with a standard previous one using simulated data. Results show that our method provides more accurate confidence intervals. This new approach is applied to the study of the extinction time of three Permo-Carboniferous synapsid taxa (Ophiacodontidae, Edaphosauridae, and Sphenacodontidae) that are thought to have disappeared toward the end of the Cisuralian, or possibly shortly thereafter. The timing of extinctions of these three taxa and of their component lineages supports the idea that a biological crisis occurred in the late Kungurian/early Roadian. -

Morphology and Evolutionary Significance of the Atlas−Axis Complex in Varanopid Synapsids
Morphology and evolutionary significance of the atlas−axis complex in varanopid synapsids NICOLÁS E. CAMPIONE and ROBERT R. REISZ Campione, N.E. and Reisz, R.R. 2011. Morphology and evolutionary significance of the atlas−axis complex in varanopid synapsids. Acta Palaeontologica Polonica 56 (4): 739–748. The atlas−axis complex has been described in few Palaeozoic taxa, with little effort being placed on examining variation of this structure within a small clade. Most varanopids, members of a clade of gracile synapsid predators, have well pre− served atlas−axes permitting detailed descriptions and examination of morphological variation. This study indicates that the size of the transverse processes on the axis and the shape of the axial neural spine vary among members of this clade. In particular, the small mycterosaurine varanopids possess small transverse processes that point posteroventrally, and the axial spine is dorsoventrally short, with a flattened dorsal margin in lateral view. The larger varanodontine varanopids have large transverse processes with a broad base, and a much taller axial spine with a rounded dorsal margin in lateral view. Based on outgroup comparisons, the morphology exhibited by the transverse processes is interpreted as derived in varanodontines, whereas the morphology of the axial spine is derived in mycterosaurines. The axial spine anatomy of Middle Permian South African varanopids is reviewed and our interpretation is consistent with the hypothesis that at least two varanopid taxa are present in South Africa, a region overwhelmingly dominated by therapsid synapsids and parareptiles. Key words: Synapsida, Varanopidae, Mycterosaurinae, Varanodontinae, atlas−axis complex, axial skeleton, Middle Permian, South Africa. -

Canine Tarsus Stabilization Surgical Technique 1 2
Canine Tarsus Stabilization Surgical Technique 1 2 Place the patient in a dorsal recumbent position and Make a medial incision beginning just proximal to the administer general anesthesia. Perform a hanging limb medial malleolus extending distally to the level of the technique with aseptic preparation and appropriate proximal intertarsal joint. Inspect the medial structures limb draping. of the tarsal joint. Identify the origin of the long part of the medial collateral ligament (MCL) on the medial malleolus of the tibia. 3 4 Using the aiming guide, place the 0.049 inch (1.2 mm) Use the 2.0 mm cannulated drill over the guidewire to guidewire from the origin of the long part of the MCL create the tibial bone tunnel for suture passage. in a proximal and slight cranial direction such that the it emerges on the lateral tibia just cranial to the fibula. Make a small incision on the lateral side where the guidewire tents the skin. 5 As a surgical landmark, identify the talocentral tarsal joint on the medial side by either a palpation or by inserting a small needle into the joint space. Using the aiming guide, place a 0.049 inch (1.2 mm) guidewire starting at the medial aspect of the talar head oriented parallel to the talocentral tarsal joint in a slightly plantar direction to capture the calcaneus. The guidewire should emerge just plantar to the boney tubercle of the lateral portion of the distal calcaneus. Make a small incision where the guidewire tents the skin. 6 7 Use the 2.0 mm cannulated drill over the guidewire to Identify the insertion of the short part of the MCL on the create the distal talus bone tunnel for suture passage. -

Curriculum Vitae
CURRICULUM VITAE AMY C. HENRICI Collection Manager Section of Vertebrate Paleontology Carnegie Museum of Natural History 4400 Forbes Avenue Pittsburgh, Pennsylvania 15213-4080, USA Phone:(412)622-1915 Email: [email protected] BACKGROUND Birthdate: 24 September 1957. Birthplace: Pittsburgh. Citizenship: USA. EDUCATION B.A. 1979, Hiram College, Ohio (Biology) M.S. 1989, University of Pittsburgh, Pennsylvania (Geology) CAREER Carnegie Museum of Natural History (CMNH) Laboratory Technician, Section of Vertebrate Paleontology, 1979 Research Assistant, Section of Vertebrate Paleontology, 1980 Curatorial Assistant, Section of Vertebrate Paleontology, 1980-1984 Scientific Preparator, Section of Paleobotany, 1985-1986 Scientific Preparator, Section of Vertebrate Paleontology, 1985-2002 Acting Collection Manager/Scientific Preparator, 2003-2004 Collection Manager, 2005-present PALEONTOLOGICAL FIELD EXPERIENCE Late Pennsylvanian through Early Permian of Colorado, New Mexico and Utah (fish, amphibians and reptiles) Early Permian of Germany, Bromacker quarry (amphibians and reptiles) Triassic of New Mexico, Coelophysis quarry (Coelophysis and other reptiles) Upper Jurassic of Colorado (mammals and herps) Tertiary of Montana, Nevada, and Wyoming (mammals and herps) Pleistocene of West Virginia (mammals and herps) Lake sediment cores and lake sediment surface samples, Wyoming (pollen and seeds) PROFESSIONAL APPOINTMENTS Associate Editor, Society of Vertebrate Paleontology, 1998-2000. Research Associate in the Science Division, New Mexico Museum of Natural History and Science, 2007-present. PROFESSIONAL ASSOCIATIONS Society of Vertebrate Paleontology Paleontological Society LECTURES and TUTORIALS (Invited and public) 1994. Middle Eocene frogs from central Wyoming: ontogeny and taphonomy. California State University, San Bernardino 1994. Mechanical preparation of vertebrate fossils. California State University, San Bernardino 1994. Mechanical preparation of vertebrate fossils. University of Chicago 2001. -

Equine MRI, Edited by Rachel C
Chapter 17 The p roximal t arsal r egion Rachel Murray , Natasha Werpy , Fabrice Audigi é , Jean - Marie Denoix , Matthew Brokken and Thorben Schulze INTRODUCTION Imaging of the proximal tarsus can be achieved in both standing and anaes- thetized horses. The height of the horse and the amount of hind end mus- culature determines which magnet confi guration will produce the best images of this region. A horse with short limbs and wide, well - muscled quarters may be more diffi cult to fi t into the long, narrow bore of some high - fi eld magnets under general anaesthesia. In the same way, the side of the lateral recumbency should be considered because it is generally easier to bring the proximal tarsus of the lower hind limb closer to the isocentre of the magnet compared with the upper one. Well developed hind limb musculature can prevent the limb from being placed far enough into the bore to allow the tarsus to be positioned at the isocentre of the magnet. The tarsus of a horse with this body type (or stature) is more easily positioned in a standing system. In contrast, for a narrow horse with long limbs, the height of the tarsus and width between the limbs may make it more diffi cult to image this region within a standing system. The degree of sway in the proximal aspect of the limbs in a standing, sedated horse is often substan- tially greater than in the distal aspect of the limbs. This degree of sway can produce signifi cant motion artefact and can make it frustrating to acquire images of the proximal tarsus. -

Physical and Environmental Drivers of Paleozoic Tetrapod Dispersal Across Pangaea
ARTICLE https://doi.org/10.1038/s41467-018-07623-x OPEN Physical and environmental drivers of Paleozoic tetrapod dispersal across Pangaea Neil Brocklehurst1,2, Emma M. Dunne3, Daniel D. Cashmore3 &Jӧrg Frӧbisch2,4 The Carboniferous and Permian were crucial intervals in the establishment of terrestrial ecosystems, which occurred alongside substantial environmental and climate changes throughout the globe, as well as the final assembly of the supercontinent of Pangaea. The fl 1234567890():,; in uence of these changes on tetrapod biogeography is highly contentious, with some authors suggesting a cosmopolitan fauna resulting from a lack of barriers, and some iden- tifying provincialism. Here we carry out a detailed historical biogeographic analysis of late Paleozoic tetrapods to study the patterns of dispersal and vicariance. A likelihood-based approach to infer ancestral areas is combined with stochastic mapping to assess rates of vicariance and dispersal. Both the late Carboniferous and the end-Guadalupian are char- acterised by a decrease in dispersal and a vicariance peak in amniotes and amphibians. The first of these shifts is attributed to orogenic activity, the second to increasing climate heterogeneity. 1 Department of Earth Sciences, University of Oxford, South Parks Road, Oxford OX1 3AN, UK. 2 Museum für Naturkunde, Leibniz-Institut für Evolutions- und Biodiversitätsforschung, Invalidenstraße 43, 10115 Berlin, Germany. 3 School of Geography, Earth and Environmental Sciences, University of Birmingham, Birmingham B15 2TT, UK. 4 Institut