Vestibular Receptors Contribute to Cortical Auditory Evoked Potentialsq
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Optogenetic Fmri Interrogation of Brain-Wide Central Vestibular Pathways
Optogenetic fMRI interrogation of brain-wide central vestibular pathways Alex T. L. Leonga,b, Yong Guc, Ying-Shing Chand, Hairong Zhenge, Celia M. Donga,b, Russell W. Chana,b, Xunda Wanga,b, Yilong Liua,b, Li Hai Tanf, and Ed X. Wua,b,d,g,1 aLaboratory of Biomedical Imaging and Signal Processing, The University of Hong Kong, Pokfulam, Hong Kong SAR, China; bDepartment of Electrical and Electronic Engineering, The University of Hong Kong, Pokfulam, Hong Kong SAR, China; cInstitute of Neuroscience, Key Laboratory of Primate Neurobiology, CAS Center for Excellence in Brain Science and Intelligence Technology, Chinese Academy of Sciences, Shanghai 200031, China; dSchool of Biomedical Sciences, Li Ka Shing Faculty of Medicine, The University of Hong Kong, Pokfulam, Hong Kong SAR, China; eShenzhen Institutes of Advanced Technology, Chinese Academy of Sciences, Shenzhen 518055, China; fCenter for Language and Brain, Shenzhen Institute of Neuroscience, Shenzhen 518057, China; and gState Key Laboratory of Pharmaceutical Biotechnology, The University of Hong Kong, Pokfulam, Hong Kong SAR, China Edited by Marcus E. Raichle, Washington University in St. Louis, St. Louis, MO, and approved March 20, 2019 (received for review July 20, 2018) Blood oxygen level-dependent functional MRI (fMRI) constitutes a multisensory integration process in the vestibular system is op- powerful neuroimaging technology to map brain-wide functions tokinetic nystagmus, whereby visual cues are used to induce in response to specific sensory or cognitive tasks. However, fMRI compensatory reflexive eye movements to maintain a stable gaze mapping of the vestibular system, which is pivotal for our sense of while moving (11, 12). These eye movements involve inputs from balance, poses significant challenges. -

Title the Human Vestibular Cortex
medRxiv preprint doi: https://doi.org/10.1101/2021.07.22.21260061; this version posted July 24, 2021. The copyright holder for this preprint (which was not certified by peer review) is the author/funder, who has granted medRxiv a license to display the preprint in perpetuity. It is made available under a CC-BY-NC 4.0 International license . Title The human vestibular cortex: functional anatomy, connectivity and the effect of vestibular disease Abbreviated title Human vestibular cortex functional anatomy Author names and affiliations Richard T. Ibitoye1,2, Emma-Jane Mallas1,3, Niall J. Bourke1, Diego Kaski4, Adolfo M. Bronstein2, David J. Sharp1,3,5 1. Computational, Cognitive and Clinical Neuroimaging Laboratory, Department of Brain Sciences, Imperial College London, London, UK. 2. Neuro-otology Unit, Department of Brain Sciences, Imperial College London, London, UK. 3. UK Dementia Research Institute, Care Research & Technology Centre, Imperial College London, London, UK 4. Department of Clinical and Motor Neurosciences, Centre for Vestibular and Behavioural Neurosciences, University College London, London, UK 5. Centre for Injury Studies, Imperial College London, London, UK Corresponding authors [email protected]; [email protected] Page/Word Counts Number of pages = 36 Number of figures = 7 Tables = 1 Number of words for Abstract = 249 Number of words for Introduction = 575 Number of words for Discussion = 1373 Conflict of interest statement The authors declare no competing financial interests. Acknowledgements This work was supported by funding from the UK Medical Research Council (MR/J004685/1), the Dunhill Medical Trust (R481/0516) and the Imperial National Institute for Health Research (NIHR) Biomedical Research Centre. -

Auditory and Vestibular Systems Objective • to Learn the Functional
Auditory and Vestibular Systems Objective • To learn the functional organization of the auditory and vestibular systems • To understand how one can use changes in auditory function following injury to localize the site of a lesion • To begin to learn the vestibular pathways, as a prelude to studying motor pathways controlling balance in a later lab. Ch 7 Key Figs: 7-1; 7-2; 7-4; 7-5 Clinical Case #2 Hearing loss and dizziness; CC4-1 Self evaluation • Be able to identify all structures listed in key terms and describe briefly their principal functions • Use neuroanatomy on the web to test your understanding ************************************************************************************** List of media F-5 Vestibular efferent connections The first order neurons of the vestibular system are bipolar cells whose cell bodies are located in the vestibular ganglion in the internal ear (NTA Fig. 7-3). The distal processes of these cells contact the receptor hair cells located within the ampulae of the semicircular canals and the utricle and saccule. The central processes of the bipolar cells constitute the vestibular portion of the vestibulocochlear (VIIIth cranial) nerve. Most of these primary vestibular afferents enter the ipsilateral brain stem inferior to the inferior cerebellar peduncle to terminate in the vestibular nuclear complex, which is located in the medulla and caudal pons. The vestibular nuclear complex (NTA Figs, 7-2, 7-3), which lies in the floor of the fourth ventricle, contains four nuclei: 1) the superior vestibular nucleus; 2) the inferior vestibular nucleus; 3) the lateral vestibular nucleus; and 4) the medial vestibular nucleus. Vestibular nuclei give rise to secondary fibers that project to the cerebellum, certain motor cranial nerve nuclei, the reticular formation, all spinal levels, and the thalamus. -
Cranial Nerve VIII
Cranial Nerve VIII Color Code Important (The Vestibulo-Cochlear Nerve) Doctors Notes Notes/Extra explanation Please view our Editing File before studying this lecture to check for any changes. Objectives At the end of the lecture, the students should be able to: ✓ List the nuclei related to vestibular and cochlear nerves in the brain stem. ✓ Describe the type and site of each nucleus. ✓ Describe the vestibular pathways and its main connections. ✓ Describe the auditory pathway and its main connections. Due to the difference of arrangement of the lecture between the girls and boys slides we will stick to the girls slides then summarize the pathway according to the boys slides. Ponto-medullary Sulcus (cerebello- pontine angle) Recall: both cranial nerves 8 and 7 emerge from the ventral surface of the brainstem at the ponto- medullary sulcus (cerebello-pontine angle) Brain – Ventral Surface Vestibulo-Cochlear (VIII) 8th Cranial Nerve o Type: Special sensory (SSA) o Conveys impulses from inner ear to nervous system. o Components: • Vestibular part: conveys impulses associated with body posture ,balance and coordination of head & eye movements. • Cochlear part: conveys impulses associated with hearing. o Vestibular & cochlear parts leave the ventral surface* of brain stem through the pontomedullary sulcus ‘at cerebellopontine angle*’ (lateral to facial nerve), run laterally in posterior cranial fossa and enter the internal acoustic meatus along with 7th (facial) nerve. *see the previous slide Auditory Pathway Only on the girls’ slides 04:14 Characteristics: o It is a multisynaptic pathway o There are several locations between medulla and the thalamus where axons may synapse and not all the fibers behave in the same manner. -
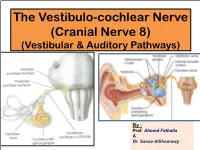
The Vestibulo-Cochlear Nerve (Cranial Nerve 8) (Vestibular & Auditory Pathways)
The Vestibulo-cochlear Nerve (Cranial Nerve 8) (Vestibular & Auditory Pathways) By : Prof. Ahmed Fathalla & Dr. Sanaa AlShaarawy OBJECTIVES At the end of the lecture, the students should be able to: qList the nuclei related to vestibular and cochlear nerves in the brain stem. qDescribe the type and site of each nucleus. qDescribe the vestibular pathways and its main connections. qDescribe the auditory pathway and its main connections. BRAIN – VENTRAL SURFACE Ponto-medullary Sulcus (cerebello- pontine angle) Vestibulo-Cochlear Nerve • Type: Special sensory (SSA) • Conveys impulses from inner ear to nervous system. • Components: § Vestibular part: conveys impulses associated with body posture ,balance and coordination of head & eye movements. § Cochlear part: conveys impulses associated with hearing. • Vestibular & cochlear parts leave the ventral surface of brain stem through the pontomedullary sulcus ‘at crebellopontine angle’ (lateral to facial nerve), run laterally in posterior cranial fossa and enter the internal acoustic meatus along with 7th nerve. Vestibular Nerve • The cell bodies (1st order neurons) are Vestibular nuclei belong to located in the vestibular ganglion special somatic afferent within the internal auditory meatus. column in brain stem. • The Peripheral processes (vestibular nerve fibers) make dendritic contact with hair cells of the membranous labyrinth (inner ear). 2 • The central processes (form the vestibular nerve) ‘’Efferent Fibres’’ : 1. Mostly end up in the lateral, medial, inferior and superior 1 vestibular nuclei (2nd order neurons) of the rostral medulla, located beneath the lateral part of the floor of 4th ventricle 2. Some fibers go to the cerebellum through the inferior cerebellar peduncle • Other Efferents from the vestibular nuclei project to other regions for the maintenance of balance, conscious awareness of vestibular stimulation , co-ordination of head & eye movements and control the posture. -

Functional Neuroanatomy for Posture and Gait Control
REVIEW ARTICLE https://doi.org/10.14802/jmd.16062 / J Mov Disord 2017;10(1):1-17 pISSN 2005-940X / eISSN 2093-4939 Functional ABSTRACT Here we argue functional neuroanatomy for pos- ture-gait control. Multi-sensory information such as Neuroanatomy somatosensory, visual and vestibular sensation act on various areas of the brain so that adaptable pos- ture-gait control can be achieved. Automatic process for Posture and of gait, which is steady-state stepping movements associating with postural reflexes including head- eye coordination accompanied by appropriate align- Gait Control ment of body segments and optimal level of pos- tural muscle tone, is mediated by the descending pathways from the brainstem to the spinal cord. Par- ticularly, reticulospinal pathways arising from the lat- Kaoru Takakusaki eral part of the mesopontine tegmentum and spinal locomotor network contribute to this process. On The Research Center for Brain Function and Medical Engineering, the other hand, walking in unfamiliar circumstance Asahikawa Medical University, Asahikawa, Japan requires cognitive process of postural control, which depends on knowledges of self-body, such as body schema and body motion in space. The cognitive in- formation is produced at the temporoparietal asso- ciation cortex, and is fundamental to sustention of vertical posture and construction of motor programs. The programs in the motor cortical areas run to ex- ecute anticipatory postural adjustment that is opti- mal for achievement of goal-directed movements. The basal ganglia and cerebellum may affect both the automatic and cognitive processes of posture- gait control through reciprocal connections with the brainstem and cerebral cortex, respectively. Conse- quently, impairments in cognitive function by dam- ages in the cerebral cortex, basal ganglia and cere- bellum may disturb posture-gait control, resulting in falling. -

The Dizzy Patient: Don't Forget Disorders of the Central Vestibular
REVIEWS The dizzy patient: don’t forget disorders of the central vestibular system Thomas Brandt1 and Marianne Dieterich1–3 Abstract | Vertigo and dizziness are among the most common complaints in neurology clinics, and they account for about 13% of the patients entering emergency units. In this Review, we focus on central vestibular disorders, which are mostly attributable to acute unilateral lesions of the bilateral vestibular circuitry in the brain. In a tertiary interdisciplinary outpatient dizziness unit, central vestibular disorders, including vestibular migraine, comprise about 25% of the established diagnoses. The signs and symptoms of these disorders can mimic those of peripheral vestibular disorders with sustained rotational vertigo. Bedside examinations, such as the head impulse test and ocular motor testing to determine spontaneous and gaze-evoked nystagmus or skew deviation, reliably differentiate central from peripheral syndromes. We also consider disorders of ‘higher vestibular functions’, which involve more than one sensory modality as well as cognitive domains (for example, orientation, spatial memory and navigation). These disorders include hemispatial neglect, the room tilt illusion, pusher syndrome, and impairment of spatial memory and navigation associated with hippocampal atrophy in cases of peripheral bilateral vestibular loss. Vertigo and dizziness are common complaints in disorders elicited by dysfunction within the temporo neuro logical patients. In a general academic emergency parietal cortex, thalamus, brainstem and cerebellum department, these symptoms were evident in 1–10% of can be classified by characteristic perceptual and ocular the overall attendances1,2, and they account for about motor manifestations, as well as sensorimotor control 13% of neurological consultations3,4. Up to 25% of of posture and gait. -

Visual Attention Modulates Glutamate-Glutamine Levels in Vestibular Cortex: Evidence from Magnetic Resonance Spectroscopy
1970 • The Journal of Neuroscience, March 3, 2021 • 41(9):1970–1981 Behavioral/Cognitive Visual Attention Modulates Glutamate-Glutamine Levels in Vestibular Cortex: Evidence from Magnetic Resonance Spectroscopy Sebastian M. Frank,1,2,3 Lisa Forster,1 Maja Pawellek,1 Wilhelm M. Malloni,1 Sinyeob Ahn,4 Peter U. Tse,2 and Mark W. Greenlee1 1Institute for Experimental Psychology, University of Regensburg, 93053 Regensburg, Germany, 2Department of Psychological and Brain Sciences, Dartmouth College, Hanover, New Hampshire 03755, 3Department of Cognitive, Linguistic, and Psychological Sciences, Brown University, Providence, Rhode Island 02912, and 4Siemens Healthcare, San Francisco, California 94123 Attending to a stimulus enhances the neuronal responses to it, while responses to nonattended stimuli are not enhanced and may even be suppressed. Although the neural mechanisms of response enhancement for attended stimuli have been intensely studied, the neural mechanisms underlying attentional suppression remain largely unknown. It is uncertain whether attention acts to suppress the processing in sensory cortical areas that would otherwise process the nonattended stimulus or the sub- cortical input to these cortical areas. Moreover, the neurochemical mechanisms inducing a reduction or suppression of neuro- nal responses to nonattended stimuli are as yet unknown. Here, we investigated how attention directed toward visual processing cross-modally acts to suppress vestibular responses in the human brain. By using functional magnetic resonance spectroscopy in a group of female and male subjects, we find that attention to visual motion downregulates in a load-depend- ent manner the concentration of excitatory neurotransmitter (glutamate and its precursor glutamine, referred to together as Glx) within the parietoinsular vestibular cortex (PIVC), a core cortical area of the vestibular system, while leaving the concen- tration of inhibitory neurotransmitter (GABA) in PIVC unchanged. -

Vestibular System
Systems Neuroscience Nov. 10, 2020 Vestibular system Daniel C. Kiper [email protected] http: www.ini.unizh.ch/~kiper/system_neurosci.html 1 VESTIBULAR SYSTEM A central role in the maintenance of equilibrium and gaze stability. The vestibular system, by means of its receptors for the perception of linear and angular acceleration, plays a central role in orientation. Designed to answer two basic questions: Which way is up? In which direction am I moving? 2 VESTIBULAR SYSTEM Very elusive to test Five peripheral “receptors” (three semicircular canals, utricule, saccule) Nerve (sub-divisions) Central connections Cortical area 3 4 The Semicircular Canals • posterior canal shares plane with contralateral anterior canal. • horizontal canals share plane. 5 Vestibular Hair Cells • Type I (aka inner) • Type II (aka outer) With Kinocilium 6 7 Responses of the Cristae • All kinocilia are oriented in the same direction • Crista in each pair of canals respond inversely to each other 8 The Otolithic Organs Saccule: roughly vertical orientation, responds to acceleration components within saggital plane Utricle: horizontal (+ 30 deg.) orientation 9 10 11 Excitation Patterns in the Utricle STRIOLA Anterior Posterior Medial 12 The otoliths register linear acceleration and static tilt 13 Vestibular system 14 15 Vestibular System 16 Vestibular Nuclei (VN) Vestibular signals originating in the two labyrinths first interact with signals from other sensory systems in the VN. Only one fraction of the neurons in the VN receive direct vestibular input, and most neurons receive afferent input from other sensory systems (visual or proprioceptive) or regions of the CNS (cerebellum, reticular formation, spinal cord and contralateral VN). -

The Vestibulo- Cochlear Nerve)
Cranial Nerve VIII (The Vestibulo- Cochlear Nerve) Lecture (11) ▪ Important ▪ Doctors Notes Please check our Editing File ▪ Notes/Extra explanation ه هذا العمل مب ين بشكل أسا يس عىل عمل دفعة 436 مع المراجعة { َوَم نْ يَ َت َو َ ّكْ عَ َلْ ا َّْلل فَهُ َوْ َحْ سْ ُ ُُْ} والتدقيق وإضافة المﻻحظات وﻻ يغ ين عن المصدر اﻷسا يس للمذاكرة ▪ Objectives At the end of the lecture, students should be able to: ✓ List the nuclei related to vestibular and cochlear nerves in the brain stem. ✓ Describe the type and site of each nucleus. ✓ Describe the vestibular pathways and its main connections. ✓ Describe the auditory pathway and its main connections. Due to the difference of arrangement of the lecture between the girls and boys slides we will stick to the girls slides then summarize the pathway according to the boys slides. Ponto-medullary Sulcus (cerebello- pontine angle) Recall: both cranial nerves 8 and 7 emerge from the ventral surface of the brainstem at the ponto- medullary sulcus Brain – Ventral Surface (cerebello-pontine angle) Vestibulo-Cochlear (VIII) 8th Cranial Nerve o Type: Special sensory (SSA) o Conveys impulses from inner ear to nervous system. o Components: • Vestibular part: conveys impulses associated with body posture ,balance and coordination of head & eye movements. • Cochlear part: conveys impulses associated with hearing. o Vestibular & cochlear parts attach to the ventral surface* of brain stem through the pontomedullary sulcus at the junction of the medulla & pons (cerebellopontine angle)* (lateral to facial nerve), run laterally in posterior cranial fossa and enter the internal acoustic meatus along with 7th (facial) nerve. -

Diagnosis of Dizziness Due to a Core Vestibular Projection Injury in a Patient with Intracerebral Hemorrhage
diagnostics Interesting Images Diagnosis of Dizziness Due to a Core Vestibular Projection Injury in a Patient with Intracerebral Hemorrhage Hyeok Gyu Kwon 1 , Chul Hoon Chang 2 and Sung Ho Jang 3,* 1 Department of Physical Therapy, College of Health Science, Eulji University, Sungnam 13135, Korea; [email protected] 2 Department of Neurosurgery, College of Medicine Yeungnam University, Daegu 42415, Korea; [email protected] 3 Department of Physical Medicine and Rehabilitation, College of Medicine, Yeungnam University, Daegu 42415, Korea * Correspondence: [email protected]; Tel.: +82-53-620-4098; Fax: +82-53-625-3508 Received: 24 March 2020; Accepted: 14 April 2020; Published: 15 April 2020 Abstract: Herein, we present a patient diagnosed with dizziness due to a core vestibular projection injury following intracerebral hemorrhage (ICH). A 51-year-old female patient underwent conservative management for a spontaneous ICH in the left hemisphere (mainly affecting the basal ganglia and insular cortex). When she visited the rehabilitation department of the university hospital at two years after the ICH onset, she advised of the presence of moderate dizziness (mainly, light-headedness) that started after ICH onset. She mentioned that her dizziness had decreased slightly over time. No abnormality was observed in the vestibular system of either ear on physical examination by an otorhinolaryngologist. However, diffusion tensor tractography results showed that the core vestibular projection in the left hemisphere was discontinued at the basal ganglia level compared with the patient’s right core vestibular projection and that of a normal subject. Therefore, it appears that the dizziness in this patient can be ascribed to a left core vestibular projection injury. -

Subjective Tests for Vestibular Dysfunction
Global Journal of Otolaryngology ISSN 2474-7556 Powerpoint presentation Glob J Otolaryngol - Special Issue March 2017 Copyright © All rights are reserved by Lalsa Shilpa Perepa DOI: 10.19080/GJO.2017.05.555664 Subjective Tests for Vestibular Dysfunction Basic Advantages a) Well established - criteria for diagnostic testing Indications pontomedullary b) Insights into site of lesion lesions produce ipsiversive tilts (deviation of a) Brandt and Dietrich [2] found that c) Alerts the examiner Pontomesencephalic lesions produce contraversive tilts subjective visual vertical toward the side of the lesion), Basicd) Disadvantages High specificity (away from the lesion). The deviations accompanied by the a) Require New and improvised versions of test ocularb) Disruption tilt reaction. of both the otolithic and vertical b) Not well supported for diagnosing semicircular canal pathways are thought to be involved inc) theThalamic deviations. lesions c) Low sensitivity may produce either ipsiversive or of the parietoinsular vestibular cortex tend to produce Subjectived) Requires Visual objective Vertical tests test to support findings contraversive tilts of subjective visual vertical. Lesions level of the thalamus and above contraversive deviations. Lesions at the Given By-Bohmer A, Rickenmann J [1]. will not produce an accompanying SVV is an estimation technique whereby a subject adjusts a oculard) Lesions tilt reaction. in the inner ear also produce deviation of visible luminescent line, while seated in complete darkness, to subjective visual vertical due to differences in the tonic whatPrinciple they consider to be upright or true vertical. outpute) Abnormal from the inotolithic headache organs in the inner ear [3]. with migraine SVV or SVH a s me a su r ed i n t he upr ig ht posit ion i s i n f luenced sufferers, particularly those byPurpose the utricles, saccules and horizontal semicircular canals.