Freshly Eclosed Chilo Partellus (Swinhoe) (Lepidoptera: Crambidae) Larvae Must Choose Whether to Feed on the Plant on Which Oviposition Occurred Or Not
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Fung Yuen SSSI & Butterfly Reserve Moth Survey 2009
Fung Yuen SSSI & Butterfly Reserve Moth Survey 2009 Fauna Conservation Department Kadoorie Farm & Botanic Garden 29 June 2010 Kadoorie Farm and Botanic Garden Publication Series: No 6 Fung Yuen SSSI & Butterfly Reserve moth survey 2009 Fung Yuen SSSI & Butterfly Reserve Moth Survey 2009 Executive Summary The objective of this survey was to generate a moth species list for the Butterfly Reserve and Site of Special Scientific Interest [SSSI] at Fung Yuen, Tai Po, Hong Kong. The survey came about following a request from Tai Po Environmental Association. Recording, using ultraviolet light sources and live traps in four sub-sites, took place on the evenings of 24 April and 16 October 2009. In total, 825 moths representing 352 species were recorded. Of the species recorded, 3 meet IUCN Red List criteria for threatened species in one of the three main categories “Critically Endangered” (one species), “Endangered” (one species) and “Vulnerable” (one species” and a further 13 species meet “Near Threatened” criteria. Twelve of the species recorded are currently only known from Hong Kong, all are within one of the four IUCN threatened or near threatened categories listed. Seven species are recorded from Hong Kong for the first time. The moth assemblages recorded are typical of human disturbed forest, feng shui woods and orchards, with a relatively low Geometridae component, and includes a small number of species normally associated with agriculture and open habitats that were found in the SSSI site. Comparisons showed that each sub-site had a substantially different assemblage of species, thus the site as a whole should retain the mosaic of micro-habitats in order to maintain the high moth species richness observed. -

Downloaded from the Same Database
G C A T T A C G G C A T genes Article Genetic Basis of Maize Resistance to Multiple Insect Pests: Integrated Genome-Wide Comparative Mapping and Candidate Gene Prioritization A. Badji 1,* , D. B. Kwemoi 2, L. Machida 3 , D. Okii 1, N. Mwila 1, S. Agbahoungba 4, F. Kumi 5 , A. Ibanda 1 , A. Bararyenya 1, M. Solemanegy 1, T. Odong 1, P. Wasswa 1, M. Otim 2, G. Asea 2, M. Ochwo-Ssemakula 1, H. Talwana 1, S. Kyamanywa 1 and P. Rubaihayo 1 1 Department of Agricultural Production, Makerere Univesity, P.O. Box 7062 Kampala, Uganda; [email protected] (D.O.); [email protected] (N.M.); [email protected] (A.I.); [email protected] (A.B.); [email protected] (M.S.); [email protected] (T.O.); [email protected] (P.W.); [email protected] (M.O.-S.); [email protected] (H.T.); [email protected] (S.K.); [email protected] (P.R.) 2 Cereals Program, National Crop Resource Research Institute, P.O. Box 7084 Kampala, Uganda; [email protected] (D.B.K.); [email protected] (M.O.); [email protected] (G.A.) 3 Alliance Bioversity International-CIAT, P.O. Box 24384 Kampala, Uganda; [email protected] 4 Laboratory of Applied Ecology, University of Abomey-Calavi, 01BP 526 Cotonou, Benin; [email protected] 5 Department of Crop Science, University of Cape Coast, P.O. Box 5007 PMB Cape Coast, Ghana; [email protected] * Correspondence: [email protected] Received: 19 May 2020; Accepted: 1 June 2020; Published: 24 June 2020 Abstract: Several species of herbivores feed on maize in field and storage setups, making the development of multiple insect resistance a critical breeding target. -

Response of Chilo Partellus (Lepidoptera: Crambidae) to Bt Maize in South Africa
Response of Chilo partellus (Lepidoptera: Crambidae) to Bt maize in South Africa J Vorster orcid.org/0000-0001-8126-6860 Dissertation submitted in fulfilment of the requirements for the Masters degree in Environmental Science at the North-West University Supervisor: Prof J van den Berg Co-supervisor: Prof MJ du Plessis Assistant supervisor: Dr A Erasmus Graduation May 2018 23441674 ACKNOWLEDGEMENTS This dissertation would not have been possible without the help of so many people. I am blessed and very grateful to have them in my life. I would like to start with our God Almighty and our Saviour who bestowed upon me the strength, wisdom and peace of mind to finish this project and who also have sent me these blessed people in my life. I would like to thank Prof. Johnnie van den Berg and Dr. Annemie Erasmus for all the guidance and support they have given me. You taught me that small things can make a big difference. Statistics can be difficult sometimes and I thank Prof. Hannalene du Plessis and Prof. Suria Elis for the help with the statistics. Thank you to all the staff at the ARC-GCI that assisted me with the trials in the lab and the planting. Elrine Strydom, Mabel du Toit, Heidi Meyer and Ursula du Plessis, thank you for the countless after hours we had to spend and for the warm hearted kindness you have given me. I would also like to thank my parents whom I dearly love for all the encouragement and motivation to do my best. You taught me that hard work does not come easily, but the fruit that you pick from it is what motivates us. -
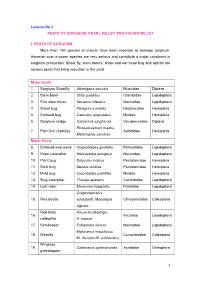
Lecture No 3 PESTS of SORGHUM, PEARL MILLET and FINGER MILLET
Lecture No 3 PESTS OF SORGHUM, PEARL MILLET AND FINGER MILLET I. PESTS OF SORGHUM More than 150 species of insects have been reported to damage sorghum. However over a dozen species are very serious and constitute a major constraint in sorghum production. Shoot fly, stem borers, shoot and ear head bug and aphids are serious pests that bring reduction in the yield. Major pests 1. Sorghum Shootfly Atherigona soccata Muscidae Diptera 2. Stem borer Chilo partellus Crambidae Lepidoptera 3. Pink stem borer Sesamia inferens Noctuidae Lepidoptera 4 Shoot bug Peregrinus maidis Delphacidae Hemiptera 5. Earhead bug Calocoris angustatus Miridae Hemiptera 6. Sorghum midge Contarinia sorghicola Cecidomyiidae Diptera Rhopalosiphum maidis, 7. Plant lice (Aphids) Aphididae Hemiptera Melanaphis sacchari Minor Pests 8. Earhead web worm Cryptoblabes gnidiella Pyraustidae Lepidoptera 9. Gram caterpillar Helicoverpa armigera Noctuidae Lepidoptera 10. Plant bug Dolycoris indicus Pentatomidae Hemiptera 11. Stink bug Nezara viridula Pentatomidae Hemiptera 12. Mirid bug Creontiades pallidifer Miridae Hemiptera 13. Slug caterpillar Thosea apierens Cochlididae Lepidoptera 14. Leaf roller Marasmia trapezalis Pyralidae Lepidoptera Cryptocephalus 15. Flea beetle schestedii, Monolepta Chrysomelidae Coleoptera signata Red hairy Amsacta albistriga, 16. Arctiidae Lepidoptera caterpillar A. moorei 17. Semilooper Eublemma silicula Noctuidae Lepidoptera Myllocerus maculosus 18. Weevils Curculionidae Coleoptera M. discolor,M. subfaciatus Wingless 19. Colemania sphenaroides Acrididae Orthoptera grasshopper MAJOR PESTS 1.Sorghum Shootfly: Atherigona soccata (Muscidae: Diptera) Distribution and status Maharashtra, Andhra Pradesh, Tamil Nadu and Karnataka Host range: Maize, ragi, bajra, rice, wheat and grasses Damage symptoms The maggot on hatching migrates to the upper surface of leaf and enters between the leaf sheath and stem. -

An Inventory of Nepal's Insects
An Inventory of Nepal's Insects Volume III (Hemiptera, Hymenoptera, Coleoptera & Diptera) V. K. Thapa An Inventory of Nepal's Insects Volume III (Hemiptera, Hymenoptera, Coleoptera& Diptera) V.K. Thapa IUCN-The World Conservation Union 2000 Published by: IUCN Nepal Copyright: 2000. IUCN Nepal The role of the Swiss Agency for Development and Cooperation (SDC) in supporting the IUCN Nepal is gratefully acknowledged. The material in this publication may be reproduced in whole or in part and in any form for education or non-profit uses, without special permission from the copyright holder, provided acknowledgement of the source is made. IUCN Nepal would appreciate receiving a copy of any publication, which uses this publication as a source. No use of this publication may be made for resale or other commercial purposes without prior written permission of IUCN Nepal. Citation: Thapa, V.K., 2000. An Inventory of Nepal's Insects, Vol. III. IUCN Nepal, Kathmandu, xi + 475 pp. Data Processing and Design: Rabin Shrestha and Kanhaiya L. Shrestha Cover Art: From left to right: Shield bug ( Poecilocoris nepalensis), June beetle (Popilla nasuta) and Ichneumon wasp (Ichneumonidae) respectively. Source: Ms. Astrid Bjornsen, Insects of Nepal's Mid Hills poster, IUCN Nepal. ISBN: 92-9144-049 -3 Available from: IUCN Nepal P.O. Box 3923 Kathmandu, Nepal IUCN Nepal Biodiversity Publication Series aims to publish scientific information on biodiversity wealth of Nepal. Publication will appear as and when information are available and ready to publish. List of publications thus far: Series 1: An Inventory of Nepal's Insects, Vol. I. Series 2: The Rattans of Nepal. -

Crambidae Biosecurity Occurrence Background Subfamilies Short Description Diagnosis
Diaphania nitidalis Chilo infuscatellus Crambidae Webworms, Grass Moths, Shoot Borers Biosecurity BIOSECURITY ALERT This Family is of Biosecurity Concern Occurrence This family occurs in Australia. Background The Crambidae is a large, diverse and ubiquitous family of moths that currently comprises 11,500 species globally, with at least half that number again undescribed. The Crambidae and the Pyralidae constitute the superfamily Pyraloidea. Crambid larvae are concealed feeders with a great diversity in feeding habits, shelter building and hosts, such as: leaf rollers, shoot borers, grass borers, leaf webbers, moss feeders, root feeders that shelter in soil tunnels, and solely aquatic life habits. Many species are economically important pests in crops and stored food products. Subfamilies Until recently, the Crambidae was treated as a subfamily under the Pyralidae (snout moths or grass moths). Now they form the superfamily Pyraloidea with the Pyralidae. The Crambidae currently consists of the following 14 subfamilies: Acentropinae Crambinae Cybalomiinae Glaphyriinae Heliothelinae Lathrotelinae Linostinae Midilinae Musotiminae Odontiinae Pyraustinae Schoenobiinae Scopariinae Spilomelinae Short Description Crambid caterpillars are generally cylindrical, with a semiprognathous head and only primary setae (Fig 1). They are often plainly coloured (Fig. 16, Fig. 19), but can be patterned with longitudinal stripes and pinacula that may give them a spotted appearance (Fig. 10, Fig. 11, Fig. 14, Fig. 22). Prolegs may be reduced in borers (Fig. 16). More detailed descriptions are provided below. This factsheet presents, firstly, diagnostic features for the Pyraloidea (Pyralidae and Crambidae) and then the Crambidae. Information and diagnostic features are then provided for crambids listed as priority biosecurity threats for northern Australia. -

Assessment of Grain Yield Losses in Pearl Millet Due to the Millet Stemborer, Coniesta Ignefusalis (Hampson)
University of Nebraska - Lincoln DigitalCommons@University of Nebraska - Lincoln Faculty Publications: Department of Entomology Entomology, Department of 2003 ASSESSMENT OF GRAIN YIELD LOSSES IN PEARL MILLET DUE TO THE MILLET STEMBORER, CONIESTA IGNEFUSALIS (HAMPSON) Aissetou Drame-Yaye Université Abdou Moumouni Ousmane Youm University of Nebraska-Lincoln, [email protected] Jonathan N. Ayertey University of Ghana Follow this and additional works at: https://digitalcommons.unl.edu/entomologyfacpub Part of the Entomology Commons Drame-Yaye, Aissetou; Youm, Ousmane; and Ayertey, Jonathan N., "ASSESSMENT OF GRAIN YIELD LOSSES IN PEARL MILLET DUE TO THE MILLET STEMBORER, CONIESTA IGNEFUSALIS (HAMPSON)" (2003). Faculty Publications: Department of Entomology. 328. https://digitalcommons.unl.edu/entomologyfacpub/328 This Article is brought to you for free and open access by the Entomology, Department of at DigitalCommons@University of Nebraska - Lincoln. It has been accepted for inclusion in Faculty Publications: Department of Entomology by an authorized administrator of DigitalCommons@University of Nebraska - Lincoln. Insect Sci. Applic. Vol. 23, No. 3,Coniesta pp. 259–265, ignefusalis 2003 larval establishment and damage 0191-9040/03 $3.00 + 0.00 259 Printed in Kenya. All rights reserved © 2003 ICIPE ASSESSMENT OF GRAIN YIELD LOSSES IN PEARL MILLET DUE TO THE MILLET STEMBORER, CONIESTA IGNEFUSALIS (HAMPSON) AISSETOU DRAME-YAYE1, OUSMANE YOUM2 AND JONATHAN N. AYERTEY3 1Faculté d’Agronomie, Université Abdou Moumouni , BP 12040 Niamey, Niger; 2 ICRISAT Sahelian Centre, BP 12404, Niamey, Niger; 3 Department of Crop Science, University of Ghana, P. O. Box LG 44, Legon Accra, Ghana (Accepted 4 June 2003) Abstract—Studies were conducted at the ICRISAT Sahelian Centre, Niger, to assess damage and yield loss by the millet stemborer, Coniesta ignefusalis (Lepidoptera: Pyralidae) on Pennisetum glaucum (L.) R. -

Biblioqraphy & Natural History
BIBLIOQRAPHY & NATURAL HISTORY Essays presented at a Conference convened in June 1964 by Thomas R. Buckman Lawrence, Kansas 1966 University of Kansas Libraries University of Kansas Publications Library Series, 27 Copyright 1966 by the University of Kansas Libraries Library of Congress Catalog Card number: 66-64215 Printed in Lawrence, Kansas, U.S.A., by the University of Kansas Printing Service. Introduction The purpose of this group of essays and formal papers is to focus attention on some aspects of bibliography in the service of natural history, and possibly to stimulate further studies which may be of mutual usefulness to biologists and historians of science, and also to librarians and museum curators. Bibli• ography is interpreted rather broadly to include botanical illustration. Further, the intent and style of the contributions reflects the occasion—a meeting of bookmen, scientists and scholars assembled not only to discuss specific examples of the uses of books and manuscripts in the natural sciences, but also to consider some other related matters in a spirit of wit and congeniality. Thus we hope in this volume, as in the conference itself, both to inform and to please. When Edwin Wolf, 2nd, Librarian of the Library Company of Phila• delphia, and then Chairman of the Rare Books Section of the Association of College and Research Libraries, asked me to plan the Section's program for its session in Lawrence, June 25-27, 1964, we agreed immediately on a theme. With few exceptions, we noted, the bibliography of natural history has received little attention in this country, and yet it is indispensable to many biologists and to historians of the natural sciences. -

Establishment of the Fungal Entomopathogen Beauveria Bassiana As an Endophyte in Sugarcane, Saccharum Officinarum
Fungal Ecology 35 (2018) 70e77 Contents lists available at ScienceDirect Fungal Ecology journal homepage: www.elsevier.com/locate/funeco Establishment of the fungal entomopathogen Beauveria bassiana as an endophyte in sugarcane, Saccharum officinarum * Trust Kasambala Donga a, b, Fernando E. Vega c, Ingeborg Klingen d, a Department of Plant Sciences, Norwegian University of Life Sciences (NMBU), Campus ÅS, Universitetstunet 3, 1433, Ås, Norway b Lilongwe University of Agriculture and Natural Resources (LUANAR), P.O. Box 219, Lilongwe, Malawi c Sustainable Perennial Crops Laboratory, United States Department of Agriculture (USDA), Agricultural Research Service, Beltsville, MD, 20705, USA d Division for Biotechnology and Plant Health, Norwegian Institute of Bioeconomy Research (NIBIO), Høgskoleveien 7, 1431, Ås, Norway article info abstract Article history: We investigated the ability of the fungal entomopathogen Beauveria bassiana strain GHA to endo- Received 18 April 2018 phytically colonize sugarcane (Saccharum officinarum) and its impact on plant growth. We used foliar Received in revised form spray, stem injection, and soil drench inoculation methods. All three inoculation methods resulted in 18 June 2018 B. bassiana colonizing sugarcane tissues. Extent of fungal colonization differed significantly with inoc- Accepted 28 June 2018 ulation method (c2 ¼ 20.112, d. f. ¼ 2, p < 0.001), and stem injection showed the highest colonization level followed by foliar spray and root drench. Extent of fungal colonization differed significantly with Corresponding Editor: James White Jr. plant part (c2 ¼ 33.072, d. f. ¼ 5, p < 0.001); stem injection resulted in B. bassiana colonization of the stem and to some extent leaves; foliar spray resulted in colonization of leaves and to some extent, the stem; Keywords: and soil drench resulted in colonization of roots and to some extent the stem. -

Interactions of Spotted Stem Borer Chilo Partellus with Wild Relatives of Sorghum
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by ICRISAT Open Access Repository Plant Breeding doi:10.1111/j.1439-0523.2012.01966.x Ó 2012 Blackwell Verlag GmbH Interactions of spotted stem borer Chilo partellus with wild relatives of sorghum 1,6 2 3 1 Venkateswaran KAMALA , Hari C. SHARMA , Daggu MANOHAR R AO , Kodeboyina S. VARAPRASAD , 4 5 Paula J. BRAMEL and Subhash CHANDRA 1National Bureau of Plant Genetic Resources, Regional Station, Rajendranagar, Hyderabad 500 030, Andhra Pradesh, India; 2International Crops Research Institute for the Semi-Arid Tropics, Patancheru 502 324, Andhra Pradesh, India; 3Department of Genetics, Osmania University, Hyderabad 500007, Andhra Pradesh, India; 4International Institute for Tropical Agriculture, Ibadan, Nigeria; 5Department of Primary Industries, Tatura 3616, Vic., Australia; 6Corresponding author, E-mail: [email protected] With 5 tables Received August 10, 2011/Accepted February 22, 2012 Communicated by T. Lu¨bberstedt Abstract owing to insect pests have been estimated to be over $1000 The levels of resistance to spotted stem borer (Chilo partellus)in million annually in the semi-arid tropics (ikisan.com 2000). sorghum are low to moderate. We therefore evaluated wild relatives of Global sorghum crop losses owing to stem borer damage have sorghum to identify accessions with high levels of resistance to this pest been estimated at over US$ 300 million annually (ICRISAT, and studied the mechanisms of resistance. Heterosorghum (Sorghum 1992; Sharma 1997). In general, yield losses range between 5% laxiflorum), Para-sorghum (S. australiense, S. purpureo-sericeum, and 10%, especially when the infestation occurs early. -

Physicochemical Mechanisms of Resistance in Sorghum to Chilo Partellus (Swinhoe)
Indian Journal of Experimental Biology Vol. 56, January 2018, pp. 29-38 Physicochemical mechanisms of resistance in sorghum to Chilo partellus (Swinhoe) Mukesh K Dhillon1* & DP Chaudhary2 1Division of Entomology, ICAR-Indian Agricultural Research Institute, New Delhi 110 012, India 2Biochemistry Laboratory, ICAR-Indian Institute of Maize Research, Pusa Campus, New Delhi 110 012, India Received 03 May 2017; revised 20 August 2017 Variation in nutritional components and amounts of secondary metabolites not only affects the growth, development and survival of insect herbivores but also indirectly influences expression of host plant resistance to insects. In this study, we examined the role of different biochemical and morphological factors in sorghum as host plant defense against the spotted stem borer, Chilo partellus (Swinhoe). The genotypes IS 2205 and IS 2123 suffered lower deadheart incidence, and exhibited deleterious effects on development and survival of C. partellus, followed by ICSV 700, ICSV 708 and ICSV 25066 than the susceptible check, Swarna. The anthocyanin pigmentation in sorghum seedlings and C. partellus deadhearts were found significantly and negatively correlated with the larval period (r = −0.60** to −0.88**), while positively correlated with the larval and pupal weights, and larval survival and adult emergence (r = 0.58* to 0.95**). Conversely, the numbers of C. partellus exit holes in the stalk, larvae recovered, number of tunnels and stem tunneling length were significantly and positively correlated (*, ** = P ≤0.05 and 0.01, respectively) with the larval period (r = 0.72** to 0.89**), but significantly and negatively correlated with larval and pupal weights, larval survival and adult emergence (r = −0.54* to −0.84**). -

Moths of the Guadalcanal Island (Lepidoptera)
Moths of the Guadalcanal Island (Lepidoptera) 著者 "KODA Nobutoyo, KUSIGEMATI Kanetosi" journal or 南方海域調査研究報告=Occasional Papers publication title volume 5 page range 87-103 URL http://hdl.handle.net/10232/15877 Kagoshima Univ. Res. Center S. Pac, Occasional Papers. No. 5. p. 87-103, 1985 87 Moths of the Guadalcanal Island (Lepidoptera) Nobutoyo K6da* and Kanetosi KUSIGEMATI** Abstract The 34 species of Heterocera of Macrolepidoptera in Guadalcanal Is. are recorded and illustrated. Introduction The butterflies of Fiji and Solomon Islands were recorded in detail by Kushi- gemati, Koda & Hirowatari (1983). The present paper deals with the remainder of the 'macrolepidoptera' which are based on material collected by the junior author in Guadalcanal Is., Solomon Islands when he joined the Scientific Expedition of Kago shima University Research Center for South Pacific, 1982. The systematic order is largely based on that of Inoue et al. (1982), and runs as follows: Pyraloidea (Hyblaeidae); Geometroidea (Uraniidae); Sphingoidea (Sphingidae); Noctuoidea (Arctiidae, Hypsidae, Ctenuchidae, Noctuidae). Information on the analysis of these species was gained mainly from references to the sources in the literatures such as Butler (1887), Druce (1888), Warren (1926), Robinson (1975) and Holloway (1979). But 7 species in the list remain unidentified, and further examinations of abundant materials are necessary in order that they may be definitely identified. Enumeration Family Hyblaeidae 1. Hyblaea onstellata Guenee, 1852 (Plate I, figs. 1, 2) Specimens examined : 2<?J< & 3^ $, 6-xii-1982, Honiara, Guadalcanal Is.; 4^ c7>, 7-xii-1982, same locality. Distribution : Indo-Australian region. This species is very common in this region. * Entomological Laboratory. Faculty of Agriculture.