EVALUATION of Deutzia X Carnea (Lem.) Rehd. AS a FOOD SOURCE to URBAN BEES
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Deutzia John Frett and Andrew Adams
Deutzia John Frett and Andrew Adams Deutzia is a large genus with more than 60 species and even more cultivars. It is a group of plants that is grown widely in the US, Europe and Asia primarily for its flowers. It has been popular in the US since its use in Victorian gardens, but the deutzia of today is nothing like that Deutzia ‘Mont Rose’ Deutzia ×kalmiiflora of days gone by. Old-fashioned Deutzia Photo: Andrew Adams Photo: Andrew Adams were more commonly large, 6–12 feet tall, upright shrubs frequently with vase Most of today’s popular Deutzia are smaller and more shape or arching habit. These plants were stunning with compact. Several of the selections offered in the sale grow typically white flowers in the spring garden, then fading 1–2 feet tall and wide, functioning more as a groundcover into the background during the summer and fall. Fruits are a than an individual shrub. These plants are best planted in dry capsule of little ornamental or wildlife value and foliage groups and are especially suitable for slopes. They are even becoming a dirty yellow before dropping in the autumn. They small enough to be integrated into the perennial border but were useful plants in larger gardens and shrub borders where do not cut them back in the fall as these shrubs flower in they could be combined with other shrubs to provide year- the spring. This means they flower on last year’s stems. If you round interest. want to tidy up these compact plants, cut them to the ground The traditional Deutzia are still after flowering and they will regrow and produce flowers very useful in today’s shrub the following spring. -

Fuzzy Pride-Of-Rochester Deutzia Scabra Thunb
Weed of the Week Fuzzy Pride-of-Rochester Deutzia scabra Thunb. Common Names: deutzia, fuzzy deutzia, pride-of- Rochester, fuzzy pride-of-Rochester Native Origin: Eastern Asia - China, Japan, Korea; introduced in 1880. Description: A large deciduous shrub in the hydrangea family (Hydrangeaceae) growing to a height 6 to 10 feet and width of 4 to 8 feet with an open, arching crown. The bark is reddish brown to orange-brown and exfoliates in large sheets. Slender stems are fuzzy/scabrous, red-brown to green, with hollow piths and pointed buds. Leaves are opposite, simple, serrate, and slightly pubescent on both surfaces, giving them a rough, fuzzy/scabrous feel. Light green leaves are 4 inches long and 2 inches wide with a rounded leaf base. Flowers are white tinged with a rosy- purple color on the outside of the corolla, fragrant, and borne on a panicle to 6 inches. Flowers are produced on the previous season’s wood and appear in late spring to early summer (April – July). The fruit is a 1/4 inch dry brown capsule that ripens in early fall and persists through the winter. It reproduces through seeds. Habitat: It grows in full sun, semi-shade, or moderately heavy shade. It tolerates most soils but prefers moist well- drained fertile soils. This ornamental plant can be found in landscaped gardens, open woodlands and shady forest edges. In Kentucky state parks fuzzy pride-of- Rochester invades woodlands areas of sugar maple, buckeye, and magnolia trees. Distribution: This species is reported from states shaded on Plants Database map. -
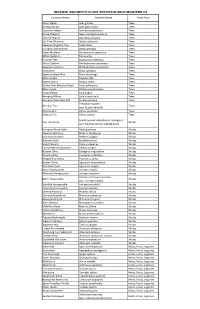
Invasive Plants in PA Web.Xlsx
INVASIVE AND WEEDY PLANT SPECIES IN SOUTHEASTERN PA Common Name Scientific Name Plant Form Amur Maple Acer ginnala Trees Norway Maple Acer platanoides Trees Sycamore Maple Acer pseudoplatanus Trees Horse Chestnut Aesculus hippocastanum Trees Tree‐of‐Heaven Ailanthus altissima Trees Silk Tree (Mimosa) Albizia julibrissin Trees Japanese Angelica Tree Aralia elata Trees European White Birch Betula pendula Trees Paper Mulberry Broussonetia papyrifera Trees White Mulberry Morus alba Trees Princess Tree Paulownia tomentosa Trees Amur Corktree Phellodendron amurense Trees Japanese Corktree Phellodendron japonicum Trees Scots Pine Pinus sylvestris Trees Japanese Black Pine Pinus thunbergii Trees White Poplar Populus alba Trees Sweet Cherry Prunus avium Trees Callery Pear (Bradford Pear) Pyrus calleryana Trees Black Locust Robinia pseudoacacia Trees Crack Willow Salix fragilis Trees Weeping Willow Salix x sepulcralis Trees European Mountain Ash Sorbus acuparia Trees Tetradium daniellii Bee‐bee Tree Trees (syn. Euodia daniellii) Chinese Elm Ulmus parvifolia Trees Siberian Elm Ulmus pumila Trees Acanthopanax sieboldianus 'variegatus' Five‐leaf Aralia Shrubs (syn. Eleytherococcus sieboldianus) European Black Alder Alnus glutinosa Shrubs Japanese Barberry Berberis thunbergii Shrubs Common Barberry Berberis vulgaris Shrubs Butterfly Bush Buddleja davidii Shrubs Scotch Broom Cytisus scoparius Shrubs Fuzzy Pride‐of‐Rochester Deutzia scabra Shrubs Russian Olive Elaeagnus angustifolia Shrubs Autumn Olive Elaeagnus umbellata Shrubs Winged Euonymus Euonymus -

Bulletin of Popular Information
ARNOLD ARBORETUM HARVARD UNIVERSITY BULLETIN OF POPULAR INFORMATION SERIES 4 VOLUME VI 1938 . VE RI I IIIII TA$ I I PUBLISHED BY THE ARNOLD ARBORETUM JAMAICA PLAIN, MASSACHUSETTS ILLUSTRATIONS Elliotia racemosa, a rare American shrub, indigenous to a restricted area in eastern and southern Georgia, Plate I, p. 11I Malus baccata mandshurica, now in full bloom, is first of the Asiatic crabapples to flower, Plate II, p. 17i The lilac path at the Arnold Arboretum, Plate III, p. 21 An airplane view of the Arnold Arboretum, Plate IV, p. 25 A fragrant double flowering Japanese cherry at the Arnold Arboretum, Plate V, p. 29 The new shade house containing the Larz Anderson Collection of dwarf trees, Plate VI, p. 33 The Larz Anderson Collection of dwarf trees now on exhibit by the Arnold Arboretum, Plate VII, p. 35 Chamaecyparis obtusa, 150 years old (Larz Anderson Collection), Plate VIII, p. 37 Robert Fortune (1813-1880), Plate IX, p. 45 Augustine Henry (1857-1930), Plate X, p. 47 Cytisus nigricans, the spike broom, Plate XI, p. 61 Acer saccharum monumentale, sentry maple, Plate XII, p. 65 Acer rubrum columnare, Plate XIII, p. 67 Views of hurricane damage in Arboretum, 1938, Plate XIV, p. 72 A stately American elm, Plate XV, p. 77i Hedge Demonstration Plot at the Arnold Arboretum, Plate XVI, pp. 82,83 iii ARNOLD ARBORETUM HARVARD UNIVERSITY BULLETIN OF POPULAR INFORMATION SERIES 4. VOL. VI APRIL 1, 1938 NUMBER l NATIONAL FLOWERS time to time we hear of the "national" flowers of various FROMEuropean countries, but it is surprising to find that there are none of these national’" flowers officially accepted by the national govern- ments. -

Plants Unlimited Slender Deutzia
[email protected] 207.594.7754 P.O. Box 374 629 Commercial St. Rockport, Maine 04856 Slender Deutzia Deutzia gracilis Height: 3 feet Spread: 4 feet Sunlight: Hardiness Zone: 5a Description: Slender Deutzia in bloom An adaptable general purpose garden shrub known for its Photo courtesy of NetPS Plant Finder frothy white flowers in spring; rather unassuming the rest of the year, use where it can be complemented by other plants when not in bloom Ornamental Features Slender Deutzia is clothed in stunning racemes of lightly-scented white trumpet-shaped flowers along the branches from mid to late spring. It has green foliage throughout the season. The serrated pointy leaves do not develop any appreciable fall color. The fruit is not ornamentally significant. Landscape Attributes Slender Deutzia is a multi-stemmed deciduous shrub with Slender Deutzia flowers a more or less rounded form. Its relatively fine texture Photo courtesy of NetPS Plant Finder sets it apart from other landscape plants with less refined foliage. This is a high maintenance shrub that will require regular care and upkeep, and should only be pruned after flowering to avoid removing any of the current season's flowers. It has no significant negative characteristics. Slender Deutzia is recommended for the following landscape applications; - Mass Planting - Hedges/Screening - General Garden Use - Container Planting Visit plants-unlimited.com [email protected] 207.594.7754 P.O. Box 374 629 Commercial St. Rockport, Maine 04856 Planting & Growing Slender Deutzia will grow to be about 3 feet tall at maturity, with a spread of 4 feet. -

FLOWERING SHRUBS Starter Plants Catalog and Shrub Reference 2018-2019 MESSAGE from DALE
FLOWERING SHRUBS Starter Plants Catalog and Shrub Reference 2018-2019 MESSAGE FROM DALE It’s just a good start. This is not negative. It’s meant to be a positive statement about how we view everything in business. It’s a way of looking at the progress that’s been made as a foundation for the future. From the latest cross made by our plant breeders to the implementation of bio-controls. From our consumer print advertising to our newest catalog. From robots sticking cuttings to building additional greenhouse space. These are really just a foundation of good things to build on. Starting with a solid foundation sets you up for unlimited positive growth and lets you take your business just about anywhere. A We’ve always included our whole staff here at the front of the catalog – now you can get to solid foundation keeps your business stable and know them all a little better on our new website. Visit springmeadownursery.com/about/meet- strong in good times and in bad. Acknowledging the-team/ or better yet, come out for a visit to see our facility and meet the team in person. that it’s merely a start and that you have to Contact your account manager for details. keep working and keep building ensures that you are constantly looking to improve and grow. As we worked on rebuilding our office after PRESIDENT PROPAGATION it was destroyed by fire in 2017, I thought a Dale Deppe, ext. 1300 Gail Berner, ext. 1404 lot about foundations. Good foundations are Jeff Flickinger, ext. -

FLORA of BEIJING Jinshuang Ma and Quanru Liu
URBAN HABITATS, VOLUME 1, NUMBER 1 • ISSN 1541-7115 FLORA OF BEIJING http://www.urbanhabitats.org Jinshuang Ma and Quanru Liu Flora of Beijing: An Overview and Suggestions for Future Research* Jinshuang Ma and Quanru Liu Brooklyn Botanic Garden, 1000 Washington Avenue, Brooklyn, New York 11225; [email protected]; [email protected] nonnative, invasive, and weed species, as well as a lst Abstract This paper reviews Flora of Beijing (He, 1992), of relevant herbarium collections. We also make especially from the perspective of the standards of suggestions for future revisions of Flora of Beijing in modern urban floras of western countries. The the areas of description and taxonomy. We geography, land-use and population patterns, and recommend more detailed categorization of species vegetation of Beijing are discussed, as well as the by origin (from native to cultivated, including plants history of Flora of Beijing. The vegetation of Beijing, introduced, escaped, and naturalized from gardens which is situated in northern China, has been and parks); by scale and scope of distribution drastically altered by human activities; as a result, it (detailing from worldwide to special or unique local is no longer characterized by the pine-oak mixed distribution); by conservation ranking (using IUCN broad-leaved deciduous forests typical of the standards, for example); by habitat; and by utilization. northern temperate region. Of the native species that Finally, regarding plant treatments, we suggest remain, the following dominate: Pinus tabuliformis, improvements in the stability of nomenclature, Quercus spp., Acer spp., Koelreuteria paniculata, descriptions of taxa, and the quality and quantity of Vitex negundo var. -

Tracheophyte of Xiao Hinggan Ling in China: an Updated Checklist
Biodiversity Data Journal 7: e32306 doi: 10.3897/BDJ.7.e32306 Taxonomic Paper Tracheophyte of Xiao Hinggan Ling in China: an updated checklist Hongfeng Wang‡§, Xueyun Dong , Yi Liu|,¶, Keping Ma | ‡ School of Forestry, Northeast Forestry University, Harbin, China § School of Food Engineering Harbin University, Harbin, China | State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing, China ¶ University of Chinese Academy of Sciences, Beijing, China Corresponding author: Hongfeng Wang ([email protected]) Academic editor: Daniele Cicuzza Received: 10 Dec 2018 | Accepted: 03 Mar 2019 | Published: 27 Mar 2019 Citation: Wang H, Dong X, Liu Y, Ma K (2019) Tracheophyte of Xiao Hinggan Ling in China: an updated checklist. Biodiversity Data Journal 7: e32306. https://doi.org/10.3897/BDJ.7.e32306 Abstract Background This paper presents an updated list of tracheophytes of Xiao Hinggan Ling. The list includes 124 families, 503 genera and 1640 species (Containing subspecific units), of which 569 species (Containing subspecific units), 56 genera and 6 families represent first published records for Xiao Hinggan Ling. The aim of the present study is to document an updated checklist by reviewing the existing literature, browsing the website of National Specimen Information Infrastructure and additional data obtained in our research over the past ten years. This paper presents an updated list of tracheophytes of Xiao Hinggan Ling. The list includes 124 families, 503 genera and 1640 species (Containing subspecific units), of which 569 species (Containing subspecific units), 56 genera and 6 families represent first published records for Xiao Hinggan Ling. The aim of the present study is to document an updated checklist by reviewing the existing literature, browsing the website of National Specimen Information Infrastructure and additional data obtained in our research over the past ten years. -

Towards a Floristic Inventory of Bat Xat Nature Reserve, Vietnam
Wulfenia 27 (2020): 233 –250 Mitteilungen des Kärntner Botanikzentrums Klagenfurt Towards a floristic inventory of Bat Xat Nature Reserve, Vietnam: Thirteen new national records of vascular plants Bui Hong Quang, Tran The Bach, Sangmi Eum, Do Van Hai, Nguyen Sinh Khang, Le Ngoc Han, Tran Duc Binh, Nguyen Thu Thuy, Vu Anh Thuong, Ngo Kien Trung, Ya-Ping Chen, Peter W. Fritsch, Chi-Ming Hu, Lu Thi Ngan, John A. N. Parnell, Alexander N. Sennikov, John R. I. Wood, Yi Yang, Andrey N. Kuznetsov, Svetlana P. Kuznetsova & Maxim S. Nuraliev Summary: The present study reports newly recorded species of vascular plants for the flora of Vietnam found in the recently established Bat Xat Nature Reserve in Lao Cai province, close to the border with Yunnan province of China. Thirteen species belonging to eleven families are reported: Acanthaceae (Strobilanthes helicta), Actinidiaceae (Actinidia melliana), Amaryllidaceae (Allium wallichii), Aquifoliaceae (Ilex fragilis), Asteraceae (Melanoseris leiolepis), Begoniaceae (Begonia yuii ), Lamiaceae (Callicarpa giraldii, Clerodendrum peii, Scutellaria macrosiphon), Lentibulariaceae (Utricularia spinomarginata), Primulaceae (Lysimachia septemfida), Pteridaceae (Aleuritopteris chrysophylla) and Symplocaceae (Symplocos glandulifera). Some of these species are additionally reported from the neighbouring Hoang Lien National Park. For each species, information on its habitat, phenology, distribution and studied specimens is provided along with the photographs of the reported findings. Keywords: Hoang Lien National Park, Indochinese Peninsula, Lao Cai province, Southeast Asia, Vietnam-China border area, Y Ty area Bat Xat Nature Reserve is located in Bat Xat district of Lao Cai province, northwestern Vietnam. It was established in 2016 by the Decision No.1954/QD-UBND of the President of Lao Cai province “On the establishment of Bat Xat Nature Reserve” (DARD 2016) in order to conserve primeval forest ecosystems in the highlands, and in particular, the rare and endangered species of flora and fauna typical of Hoang Lien Son mountain region. -
Flowering Shrubs
WATERDOWN 905.689.7433 FLOWERING SHRUBS TRENTON 613.392.0402 www.connon.ca • [email protected] COTONEASTER DEUTZIA ACUTIFOLIUS PEKING COTONEASTER Code: 5570 Height: 3m Spread: 3m Zone: 4 Shape: Upright Foliage is dull green and hairy. Black fruit. Excellent plant for hedging. Fall colour ranges from yellow-red-purple. DEUTZIA CRÉME FRAICHE COTINUS COGGYGRIA GRACE CRÈME FRAICHE CRÈME FRAICHE DEUTZIA COGGYGRIA GRACE Code: 5371 Height: 60cm Spread: 60cm GRACE SMOKEBUSH Zone: 5 Shape: Round Code: 5550 Height: 6m Spread: 6m Star-shaped pure white flowers Zone: 4 Shape: Round appear in the Spring with variegated Soft purple-red leaves. Purple-pink foliage. Well suited for small spaces flowers in summer turn to a beautiful COTONEASTER APICULATUS or as a groundcover. Deer resistant. scarlet colour in the fall. APICULATUS CRANBERRY COTONEASTER Code: 5590 Height: 1m Spread: 2m Zone: 4 Shape: Spreading Low, wide-spreading shrub with stiff branching. Cranberry-red fruit in August-September. APICULATUS TOM THUMB TOM THUMB CRANBERRY COTONEASTER Code: 5600 Height: 1m Spread: 2m Zone: 4 Shape: Spreading COTINUS COGGYGRIA ROYAL PURPLE Neat, mounding shrub with charming DEUTZIA CRENATA NIKKO COGGYGRIA pink flowers followed by bright red ROYAL PURPLE bead-like berries. CRENATA NIKKO ROYAL PURPLE SMOKEBUSH NIKKO SLENDER DEUTZIA Code: 5660 Height: 60cm Spread: 1.5m Code: 5565 Height: 4m Spread: 4m CRATAEGUS Zone: 5 Shape: Upright Zone: 5 Shape: Spreading Leaves emerge rich maroon-red and CRUS-GALI CLUMP A compact, spreading groundcover darken to reddish-black. Rich purple WASHINGTON that produces an abundance fall colour. of snowy white flowers in May. HAWTHORN CLUMPFORM Maroon fall colour. -

Chalet Nursery Nikko Deutzia
Nikko Deutzia Deutzia gracilis 'Nikko' Height: 3 feet Spread: 4 feet Sunlight: Hardiness Zone: 4 Other Names: Slender Deutzia Description: Nikko Deutzia in bloom A refined garden detail plant, valued for its low mounded Photo courtesy of NetPS Plant Finder habit, frothy white flowers in spring and good fall color; great for massing or use in the front of a shrub border Ornamental Features Nikko Deutzia is smothered in stunning racemes of lightly-scented white trumpet-shaped flowers along the branches from mid to late spring. It has green foliage throughout the season. The small serrated pointy leaves turn an outstanding deep purple in the fall. The fruit is not ornamentally significant. Landscape Attributes Nikko Deutzia is a multi-stemmed deciduous shrub with a more or less rounded form. Its relatively fine texture sets it apart from other landscape plants with less refined foliage. This is a relatively low maintenance shrub, and should only be pruned after flowering to avoid removing any of the current season's flowers. It has no significant negative Nikko Deutzia flowers characteristics. Photo courtesy of NetPS Plant Finder Nikko Deutzia is recommended for the following landscape applications; - Mass Planting - General Garden Use - Groundcover - Container Planting 3132 LAKE AVENUE W ILM ETTE, ILLINOIS 60091 847.256.0561 W W W.CHALETNURSERY.COM Planting & Growing Nikko Deutzia will grow to be about 3 feet tall at maturity, with a spread of 4 feet. It tends to fill out right to the ground and therefore doesn't necessarily require facer plants in front. It grows at a slow rate, and under ideal conditions can be expected to live for approximately 20 years. -

Wilson's Controversial Magnolia and the Mysterious Wa Shan
photograph © President and Fellows of Harvard College. Arnold Arboretum Archives. Wilson’s photo of Wa Shan. “The fog prevented my view of the wild and savage scenery for which this stretch is famed. I was very disappointed as I was keen on getting some photos” (Wilson’s diary 18 September, 1908); “the Mt. covered in dense cloud all day” (19 September); “Mt. Wa was clear, 52 and I took 3 photos from different positions” (20 September). Wilson’s controversial magnolia and the mysterious Wa Shan As is so often the case, what was expected to be a leisurely pursuit of magnolia study in the tranquil atmosphere of the Australian Bicentennial Arboretum, turned out to be an exciting adventure into the wilderness of China’s remote and elusive Wa Shan, a mountain shrouded in mist and mystery. This is such a tale when CHRIS CALLAGHAN and SK PNG went searching for Wilson’s controversial Magnolia sargentiana var. robusta in a quest to shed further light on its botanical status and ended up rediscovering a mountain potentially richer in flora than Emei Shan! While researching the identity of what was found to be probably the first recorded hybrid of Magnolia sargentiana var. sargentiana (see ‘Bicentennial’ – a new hybrid magnolia cultivar, in the 2008 IDS Yearbook, pp. 53 - 61), I became aware of a controversy surrounding the botanical status of Magnolia sargentiana var. robusta, the few plants of which were discovered by Ernest Wilson in woodlands and open country on Wa Shan in Sichuan province, INTERNATIONAL DENDROLOGY SOCIETY TREES photograph © Australian Bicentennial Arboretum Da-tien Chi at base of Wa Shan in October 2009.