Description and Taphonomic Investigations of Neogene Proboscidea from Rhodos, Greece*
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Monolithos Castle, Rhodes Island, Greece
Monolithos Castle, Rhodes island, Greece Monolithos Castle at the island of Rhodes in Greece Monolithos Castle is situated on Rhodes island, the largest of the Dodecanese islands located near the Aegean sea. The exotic location of the Rhodes island between the Greek mainland and the island of Cyprus makes it one of the popular tourist destinations in the world. Apart from these, there are a number of sites of historical and archeological importance that attract tourists. The Monolithos Castle is definitely one of the most popular places for sightseeing in Rhodes. The Monolithos Castle offers scenic views of the coastline. The location of the castle makes it a perfect place to bask in the bounties of nature. Climbing the rock to reach the castle is not as difficult as it appears. You need to climb some steps that pass through the lush green area. But, there are a few slippery steps, so you need to be careful. Even though the castle is small, yet if you have a close look, you will find that here are two hidden rooms. From the top you will be able to see courtyards dabbed in whitewash and decorated with geraniums, isolated beaches where people play games and splash in the waves, and stone houses with tiled roofs overlooking the great expanse of azure blue water that splashes upon the beach. These are the views of the nearby village that appears like an amphitheater. There are two small 15th century churches nearby, dedicated to Saint Agios Panteleimon and Saint Geroge. Both chapels are in ruins today. -

The Herpetofauna of the Island of Rhodes (Dodecanese, Greece)
HERPETOZOA 21 (3/4): 147 - 169 147 Wien, 30. Jänner 2009 The herpetofauna of the Island of Rhodes (Dodecanese, Greece) Die Herpetofauna der Insel Rhodos (Dodekanes, Griechenland) THOMAS BADER & CHRISTOPH RIEGLER & HEINZ GRILLITSCH KURZFASSUNG Die Herpetofauna von Rhodos wird aufgrund von Literaturdaten, Sammlungsmaterial des Naturhistorischen Museums in Wien sowie Ergebnissen diverser herpetologischer Exkursionen aufgelistet. Verbreitungskarten aller nachgewiesenen Arten sowie eine aktuelle herpetologische Faunenliste werden erstellt. Folgende Arten wurden regelmäßig und wiederholt festgestellt: Bufo viridis, Hyla arborea, Pelophylax cerigensis, Mauremys rivulata, Hemidactylus turcicus, Laudakia stellio, Ophisops elegans, Lacerta trilineata, Anatolacerta oertzeni, Ablepharus kitaibelii, Chalcides ocellatus, Trachylepis aurata, Blanus strauchi, Typhlops vermicularis, Dolichophis sp., Platyceps najadum, Hemorrhois nummifer, Zamenis situla, Natrix natrix, Telescopus fallax. Der taxonomische Status der Springnatter wird diskutiert. Aufgrund mehrerer aktueller Funde wird Mediodactylus kotschyi ergän- zend zur Herpetofauna von Rhodos hinzugefügt und das Auftreten folgender Arten, deren Vorkommen in der Literatur regelmäßig für Rhodos angegeben wurde, in Frage gestellt: Testudo graeca, Pseudopus apodus, Natrix tessellata. Aktuelle Einzelfunde gelangen von Chamaeleo chamaeleon, Malpolon monspessulanus, Testudo grae- ca und Testudo hermanni, deren Status diskutiert wird. ABSTRACT Based on several recent herpetological surveys, as well as data from -

First Annotated Checklist of Chironomidae of Rhodos, Greece
SPIXIANA 31 2 223–231 München, November 2008 ISSN 0341–8391 First annotated checklist of Chironomidae of Rhodos, Greece (Insecta, Diptera) Arnold Móra & Zoltán Csabai Móra, A. & Z. Csabai (2008): First annotated checklist of Chironomidae of Rhodos, Greece (Insecta, Diptera). – Spixiana 31/2: 223-231 Chironomid fauna of Greek Aegean islands, an essential part of the biogeo- graphically important Mediterranean region, is almost unexplored, with only 36 species recorded prior to the present study. It is especially true for Dodecanese islands (6 recorded species). In 2007, chironomid larvae and exuviae were collected in Rhodos, the largest member of the Dodecanese islands. Thirty-four taxa were identified (8 Tanypodinae, 10 Orthocladiinae, 16 Chironominae). None of the pre- viously recorded taxa were found; thus all collected species proved to be new for the fauna of Rhodos. An actualized checklist for the Chironomidae of Rhodos is given based on the literature and results of recent investigations. Keywords: Dodecanese islands, distribution, larvae, pupal exuviae Arnold Móra, Hungarian Academy of Sciences, Balaton Limnological Research Institute, Klebelsberg Kuno 3., H-8237 Tihany, Hungary; e-mail: marnold@tres. blki.hu Zoltán Csabai, University of Pécs, Department of General and Applied Ecology, Ifjuság útja 6., H-7624 Pécs, Hungary; e-mail: [email protected] Introduction macroinvertebrate fauna of Rhodos, the largest of the Dodecanese islands. During this expedition chirono- As regards the Chironomidae, the Mediterranean mid larvae and pupal exuviae were also collected. region constitutes a link between the Palaearctic The aims of this paper are to present new faunistical and Afrotropical, and maybe the Oriental biogeo- data on the basis of those recent collections, and to graphical regions (Laville & Reiss 1992). -

Περίληψη : Γενικές Πληροφορίες Area: 1,401.46 Km2
IΔΡΥΜA ΜΕΙΖΟΝΟΣ ΕΛΛΗΝΙΣΜΟΥ Συγγραφή : Παπαχριστοδούλου Ιωάννης , Παλαιοθόδωρος Δημήτρης , Παλαιοθόδωρος Δημήτρης , Βαξεβάνης Γιάννης , Σπυροπούλου Βάσω , Βαξεβάνης Γιάννης , Μπαζίνη Ελένη , Πατσιαδά Βασιλική , Σέλελη Περίληψη : Γενικές Πληροφορίες Area: 1,401.46 km2 Coastline length: 253 km Population: 117,007 Island capital and its population: Rhodes (52,318) Administrative structure: Region of South Aegean, Prefecture of Dodecanese, Municipality of Rhodes (Capital: Rhodes, 52,318), Municipality of Archangelos (Capital: Archangelos, 5,500), Municipality of Attavyros (Capital: Empona, 1,216), Municipality of Afandou (Capital: Afandou, 5,494), Municipality of Ialysos (Capital: Trianta, 10,107 people), Municipality of Lindos (Capital: Lindos, 810), Municipality of Kalithea (Capital: Faliraki, 1,491), Municipality of Kamiros (Capital: Soroni, 1,236), Municipality of North Rhodes (Capital: Gennadi, 655), Municipality of Petaloudes (Capital: Kremasti, 4,372). Local newspapers: Proodos, Rodiaki. Dodekanisos, Gnomi, Drasis, Dimokratiki tis Rodou, Angelia, Vima tis Dodekanisou. Local Radio station: Radio Anatoli (87.5), Radio 1 (88.0), Super Star (89.1), Radio Enosi (89.6), Christianismos (90.0), Rodos 90.7 (90.7), Radio Lichnari (91.4 and 91.7), G.R.Α. Rodos (92.7 και 98.4), Rythmos FM (93.5), Antenna FM (93.8 και 94.4), Radio Archangelos (94.1 and 101.6), SGL Radio (95.1), FM1 (95.6), Nei Orizontes (95.9), Radio Methorios (96.9), Lemon Radio (97.3), Alpha Radio Dodekanisou (97.7), Love NRG (98.0), Appolon FM (98.4), Sky Rhodes 100 (99.0), Palmos -

1. Introduction 2. the Island of Rhodes
WATER SUPPLY MANAGEMENT APPROACHES USING RES IN RHODES ISLAND, GREECE E. Manoli, D. Assimacopoulos1 and C. A. Karavitis2 National Technical University of Athens, Chemical Engineering Department, Process Analysis and Plant Design Section, 9 Heroon Polytechniou, Zografou Campus, GR-15780, Athens, Greece ABSTRACT: Desalination powered by Renewable Energy Sources (RES) is presented in order to augment the water supply in the semi-arid region of Rhodes island.. The island of Rhodes was chosen as a case study as it relies mostly on the exploitation of groundwater resources and faces serious water shortage problems due to the aquifers depletion. Alternatives are discussed and compared in contrast to the construction of storage dams so as to meet the urban demand up to the year 2040. Results may indicate that by using also financial incentives and management approaches, desalination powered by RES could be an attractive and environmentally friendly option in an effort to solve problems related to water quantity and quality in semi-arid regions with adequate Renewable Energy potential. 1. INTRODUCTION resources problems. The whole scheme should be incorporated in an overall management framework in an In the South Mediterranean region water is used in an effort to ensure the long term sustainability of the region. unsustainable manner. The South European Mediterranean landscape, as a whole, is ecologically fragile and seriously endangered by prevailing social and economic trends. In 2. THE ISLAND OF RHODES this regard, the future of the region may be threatened by increasing coastal area stress, by expanding differences 2.1 Physiography, Social and Economic Profile between tourist areas and the rural hinterlands, and by the sensitivity between the water and soil equilibrium. -

Voyage Naturaliste À Rhodes (7 Au 27 Avril 2003)
Voyage naturaliste à Rhodes (7 au 27 avril 2003) Georges et Mireille Olioso, le Mail, 26230 Grignan (France) Considérations générales Il y a un important contraste entre la partie nord de l’île, avec une forte implantation touristique, et la partie sud, beaucoup moins peuplée, mais cela ne saurait durer vu le rythme des constructions. En cette saison, les touristes ne sont pas encore très nombreux et se cantonnent dans les sites historiques. La circulation est facile mais il convient de rester vigilent car, sur les deux grands axes côtiers, le code de la route (particulièrement les lignes blanches) n’est pas très bien respecté (c’est un euphémisme !). La nuit, c’est pire, attention aux rares feux tricolores vers 6h du matin, certains font comme s’ils n’existaient pas ! Quant aux limitations de vitesse … quand vous lisez 50 km/h, la plupart doivent lire 80 km/h… Les routes sont assez bien entretenues bien qu’on y rencontre parfois des trous de belle taille. L’essence sans plomb coûte entre 0,73 et 0,81 €, le diesel entre 0,69 et 0,75 €. Il y a de nombreux hôtels ouverts et des restaurants un peu partout, ouverts surtout le soir. Au point de vue ornithologique, nous sommes probablement tombés dans une situation météorologique des plus favorables qui nous a permis d’observer un grand nombre de migrateurs, certains très rares. Il convient de visiter en particulier toutes les embouchures de fleuves (il y en a pas mal), surtout sur la côte est. C’est cette côte qui voit arriver les migrateurs en provenance d’Afrique, et plus particulièrement la région la plus méridionale. -

Pliocene Vertebrate Locality of Qalta, Ankara, Turkey. 12. Paleoecology, Biogeography and Biochronology
Pliocene vertebrate locality of Qalta, Ankara, Turkey. 12. Paleoecology, biogeography and biochronology Sevket SEN Laboratoire de Paléontologie, URA 12 du CNRS, Muséum national d'Histoire naturelle, 8 rue de Buffon, F-75231 Paris cedex 05 (France) [email protected] Geneviève BOUVRAIN Laboratoire de Paléontologie des Vertébrés, URA 1761 du CNRS, université Pierre-et-Marie-Curie, 4 place Jussieu, F-75252 Paris cedex 05 (France) Denis GERAADS EP 1781 du CNRS, Collège de France, 30 avenue Marcelin-Berthelot, F-92360 Meudon (France) Sen S., Bouvrain G. & Geraads D. 1998. — Paleoecology, biogeography and biochrono logy, in Sen S. (ed.), Pliocene vertebrate locality of Qalta, Ankara, Turkey. 12, Geodiversitas 20 (3) : 497-510. ABSTRACT Integrated studies on the Pliocene vertebrate locality of Calta provided abun dant data on the environment of the fauna, its biogeographic relationships and its age. The fauna is dominated by animals adapted to relatively dry and KEYWORDS open environments with loose soil and some watet points. The presence of Mammals, short-limbed herbivores and of ochotonids might indicate a landscape with Pliocene, a ta Ruscinian, some relief nearby. The C l fauna is mainly composed of elements with Turkey, European and some Asiatic affinities. African affinities of some taxa are paleoenvironment, doubtful. The time range of some mammalian groups is discussed, the Caha biogeography, biochronology. locality is dated as eatly MN15 (late Ruscinian). 1998 • 20 (3) 497 Sen S., Bouvrain G. & Geraads D. RÉSUMÉ Le gisement de vertébrés pliocenes de Çalta, Ankara, Turquie. 12. biogéographie et biochronologie. Des recherches multidisciplinaires sur la faune de vertébrés et les sédiments de Çalta ont permis d'obtenir des résultats sur l'environnement, les affinités de la faune et son âge. -
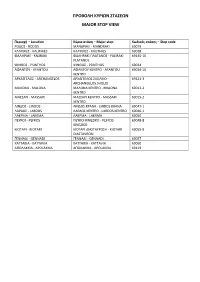
Προβολη Κυριων Στασεων Major Stop View
ΠΡΟΒΟΛΗ ΚΥΡΙΩΝ ΣΤΑΣΕΩΝ MAJOR STOP VIEW Περιοχή – Location Κύρια στάση – Major stop Κωδικός στάσης – Stop code ΡΟΔΟΣ - RODOS ΜΑΝΔΡΑΚΙ - MANDRAKI 69079 ΚΑΛΥΘΙΕΣ - KALITHIES ΚΑΛΥΘΙΕΣ - KALITHIES 69028 ΦΑΛΗΡΑΚΙ - FALIRAKI ΦΑΛΗΡΑΚΙ ΠΛΑΤΑΝΟΣ - FALIRAKI 69145-10 PLATANOS ΨΙΝΘΟΣ - PSINTHOS ΨΙΝΘΟΣ - PSINTHOS 69034 ΑΦΑΝΤΟΥ - AFANTOU ΑΦΑΝΤΟΥ ΚΕΝΤΡΟ - AFANTOU 69024-10 KENTRO ΑΡΧΑΓΓΕΛΟΣ - ARCHANGELOS ΑΡΧΑΓΓΕΛΟΣ ΣΧΟΛΕΙΟ - 69121-3 ARCHANGELOS SHOLIO ΜΑΛΩΝΑ - MALONA ΜΑΛΩΝΑ ΚΕΝΤΡΟ - MALONA 69013-2 KENTRO ΜΑΣΣΑΡΙ - MASSARI ΜΑΣΣΑΡΙ ΚΕΝΤΡΟ - MASSARI 69015-2 KENTRO ΛΙΝΔΟΣ - LINDOS ΛΙΝΔΟΣ ΚΡΑΝΑ - LINDOS KRANA 69047-1 ΛΑΡΔΟΣ - LARDOS ΛΑΡΔΟΣ ΚΕΝΤΡΟ - LARDOS KENTRO 69046-1 ΛΑΕΡΜΑ - LAERMA ΛΑΕΡΜΑ - LAERMA 69050 ΠΕΥΚΟΙ - PEFKOS ΠΕΥΚΟΙ ΚΙΝΕΖΙΚΟ - PEFKOS 69048-8 KINEZIKO ΚΙΟΤΑΡΙ - KIOTARI ΚΙΟΤΑΡΙ ΔΙΑΣΤΑΥΡΩΣΗ - KIOTARI 69055-9 DIASTAVROSI ΓΕΝΝΑΔΙ - GENNADI ΓΕΝΝΑΔΙ - GENNADI 69057 ΚΑΤΤΑΒΙΑ - KATTAVIA ΚΑΤΤΑΒΙΑ - KATTAVIA 69060 ΑΠΟΛΑΚΚΙΑ - APOLAKKIA ΑΠΟΛΑΚΚΙΑ - APOLAKKIA 69119 ΑΝΑΛΥΤΙΚΗ ΠΡΟΒΟΛΗ ΣΤΑΣΕΩΝ DETAILED STOP VIEW Κωδικός στάσης – Stop code Όνομα στάσης Stop name 69028 ΚΑΛΥΘΙΕΣ KALITHIES 69034 ΨΙΝΘΟΣ PSINTHOS 69050 ΛΑΕΡΜΑ LAERMA 69051 ΠΥΛΩΝΑ PYLONA 69056 ΒΑΤΙ VATI 69057 ΓΕΝΝΑΔΙ GENNADI 69060 ΚΑΤΤΑΒΙΑ KATTAVIA 69063 ΛΑΧΑΝΙΑ LAHANIA 69067 ΠΡΟΦΙΛΙΑ PROFILIA 69079 ΜΑΝΔΡΑΚΙ MANDRAKI 69119 ΑΠΟΛΑΚΚΙΑ APOLAKKIA 69120 ΑΡΝΙΘΑ ARNITHA 69122 ΑΣΚΛΗΠΙΕΙΟ ASKLIPIO 69131 ΙΣΤΡΙΟ ISTRIO 69144 ΤΣΑΜΠΙΚΑ TSAMPIKA 69147 Χ.ΜΑΝΩΛΗ H.MANOLI 69147-1 Χ.ΜΑΝΩΛΗ Ε. H.MANOLI E. 69079-1 ΠΑΡΚΟ ΔΗΜΟΚΡΑΤΙΑΣ PARKO DIMOKRATIAS 69079-2 ΠΑΝΕΠΙΣΤΗΜΙΟ UNIVERSITY 69079-3 ΣΑΝ ΦΡΑΤΖΙΣΚΟ SAN FRANCISCO 69079-11 -

Rhodes (Rhodos): 4-11 April 2007
Rhodes (Rhodos): 4-11 April 2007 I went to the Greek island of Rhodes (Rhodos) during the week from 4 to 11 April 2007. It was just me and my girlfriend Luisa, and we took a rather relaxed holiday, surely not a 6 am – 8 pm birding thing! Before going, I checked the available web resources, but I did not purchase any specific book, apart from a 1:100.000 map (Road Editions, nr. 202). It was clear that there are not dozens of report or other useful infos, in fact Rhodes seems to be pretty ignored by most birders. I was in touch with Jos Rutten and I even met him twice during our stay (the first one... by chance!). He's from Belgium and he seems to be the most expert birder for that island; he gave me an interesting Rhodes check list which turned out to be very useful. In my small web site there's a gallery of sites and some birds mentioned in this report. I also had, but actually never used, a brochure from www.rodosisland.gr which contains information on many walking trails on the island, including small photos and maps; the pages were originally by Graf Editions (2004); this document anyway is not at all about birdwatching. We stayed in Sun Beach hotel which is in the Ialysos area, in the NW side, between the airport and Rhodes town. Actually I suggest to choose an accommodation in the opposite side of the island, which is more or less just north or south of Lindos. -

Adventure Tours Europe #B1/2407 Rhodes Island 7-Day Adventure Tour
Full Itinerary and Tour details for Rhodes Island 7-day Adventure Tour Prices starting from. Trip Duration. Max Passengers. 1125 € 7 days 16 Country. Greece Experience. Tour Code. Adventure Tours Europe #B1/2407 Rhodes Island 7-day Adventure Tour Tour Details and Description Explore the beauty of a destination and make new friends on this 7-day stay with a selection of activities:horse-riding, scuba diving, hiking, cycling and discovering nature - a great experience in ecotourism.The guest house is part of the facilities in the adventure park of Apolakkia lake in southern Rhodes. The wooden guest house is simple and always clean. It has 7 beds, 3 toilets and 3 showers. The zone is protected by the European Union (Natura 2000) and the landscape is ideal for those seeking to enjoy the peace and quiet and the beauty of nature. Live in the heart of nature and enjoy daily activities. Included in the Rhodes Island Tour: 7-day/6-night stay in our Guest house at the lake of Apolakkia 4-day tour with activities – Horse riding, Scuba diving, Climbing, Sailing – and the hot spots of the island. Activities around the lake where the guest house is located -Archery, Canoe, Flying fox, Biking & Hiking. Daily breakfast & lunch Transfer from/to the airport Van support from convenient locations in southern Rhodes Insurance for tours & activities Equipment & lessons for the activities English / German / French speaking guides with extensive local knowledge More options for activities and tours (e.g. yoga, agritourism courses, rhodes town nightlife tour) Check Availability Book Online Now Send an Enquiry 2/6 Rhodes Island 7-day Adventure Tour Daily Tours & Activities: Horse-riding, Archery & southern Rhodes treasures. -

Efstratios Koufos
CV EFSTRATIOS KOUFOS Efstratios Koufos Rhodes island, Greece Phone: +30 693 2079623 Email: [email protected] Web: http://stratos-photography.wix.com/stratosjmj Education University of Aegean, Life Long Learning Programme (2013-2014) Courses attended: Data bases and use of GIS in Archaeoastronomy Introduction to Archaeoastronomy Archaeological research and interpretation Astronomical alignments of monuments Basic statistical methods Lyceum of Rhodes (Graduated 1989) Skills & Professional Experience A. Excavation Specialist, Ephorate of Antiquities, Dodecanese, Greece Most Important Excavations (Rhodes) 2014-2015: Excavation of the prehistoric cemetery in the region “Agathi” of the city of Haraki. 1998-2013: Excavation of large part of the Hellenistic wall of the “Jetty of Mills” 1994-1998: Excavation of the Hellenistic depository in the surrounding area of St. Catherine’s hostel. 1989-1994: Excavation of south collashium of nights at the Castello (complex of the knights’ houses) and discovery of the central temple of “St. John of the Knights”. Other On-going Excavations (Rhodes) Large scale tubes of Prehistoric, Hellenistic and Medieval periods distributed throughout the complex of the medieval city of Rhodes. Rescue excavations in monuments under restoration Trial excavations throughout the island of Rhodes Trial excavations with important discoveries of the older parts of the fortification of the medieval city of Rhodes and of the corresponding moat CV: Efstratios Koufos Extensive Experience with: Emergency repairs, maintenance reconstruction and restoration works of monuments First degree restoration of antiquities Transportation of mosaics from the excavation site Organization of the byzantine museum at the Castello of Rhodes Underwater photography of byzantine shipwreck for the Ephorate of underwater antiquities Organization of technical specifications at excavation sites (collaboration w/ archaeologists) Measurements of levels and archaeological layers with leveler Top view photography of excavation sites and related photomerge B. -

Rhodes Island
RHODES ISLAND Rhodes town is the largest settlement and capital city of the island. It is famous as the ancient site of the Colossus (c 280BC), one of the seven wonders of the ancient world, and as the oldest inhabited medieval town in Europe. Rhodes was declared a World Heritage City by UNESCO in 1988. Now about 6000 people live and work in the same buildings the Knights of St. John lived in six centuries ago. The total population of the city is roughly 100 000. Rhodes is one of the largest and most fertile of the Greek Islands, and is one of the most visited because of its combination of beaches, archaeological sites, and extensive medieval town. The climate is particularly good, with the weather typically sunny and mild. The island is usually counted as one of the Dodecanese, but due to its importance for travelers is considered separately here. The rock-rose is so prolific here that it has been named the 'Island of Roses,' though modern scholars doubt the ancient theory that the island's name comes from the Greek word for rose. While the northern coast is renowned for its lively tourist resorts the south offers tranquil beaches and a slower, simpler pace of life. Harbour gates in Rhodes city The Colossus of Rhodes was a statue of the Greek titan-god of the sun Helios, erected in the city of Rhodes, on the Greek island of the same name, by Chares of Lindos in 280 BC. One of the Seven Wonders of the Ancient World, it was constructed to celebrate Rhodes' victory over the ruler of Cyprus, Antigonus I Monophthalmus, whose son unsuccessfully besieged Rhodes in 305 BC.