The Distribution of Oceanic Cirripedes in the North-East Atlantic in Summer 1983 and the Connotations of the Results to the Problems of Conchoderma Fouling
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

(Cirripedia : Thoracica) Over the Body of a Sea Snake, Laticauda Title Semifasciata (Reinwardt), from the Kii Peninsula, Southwestern Japan
Distribution of Two Species of Conchoderma (Cirripedia : Thoracica) over the Body of a Sea Snake, Laticauda Title semifasciata (Reinwardt), from the Kii Peninsula, Southwestern Japan Yamato, Shigeyuki; Yusa, Yoichi; Tanase, Hidetomo; Tanase, Author(s) Hidetomo PUBLICATIONS OF THE SETO MARINE BIOLOGICAL Citation LABORATORY (1996), 37(3-6): 337-343 Issue Date 1996-12-25 URL http://hdl.handle.net/2433/176259 Right Type Departmental Bulletin Paper Textversion publisher Kyoto University Pub!. Seto Mar. Bioi. Lab., 37(3/6): 337-343, 1996 337 Distribution of Two Species of Conchoderma (Cirripedia: Thoracica) over the Body of a Sea Snake, Laticauda semifasciata (Reinwardt), from the Kii Peninsula, Southwestern Japan SHIGEYUKI YAMATO, YOICHI YUSA and HIDETOMO TANASE Seto Marine Biological Laboratory, Kyoto University, Shirahama, Wakayama 649-22, Japan Abstract Two species of Conchoderma were found on a sea snake, Laticauda semifas ciata (Reinwardt), collected on the west coast of the Kii Peninsula. A total of 223 individuals of C. virgatum and 6 of C. hunteri in 19 clumps were attached to the snake's body. The barnacles ranged in size from 1.4 mm (cypris larvae) to 18.2 mm in capitulum length in C. virgatum, and from 10.7 to 14.4 mm in C. hunteri. The size of the smallest gravid individuals in both species was between 10 and 11 mm. The distribution of C. virgatum on the snake was non-random both longitudinally and dorso-ventrally, with more barnacles in the posterior region and on the ventral side of the snake, respectively. The proportion of gravid individuals increased towards the tail. -
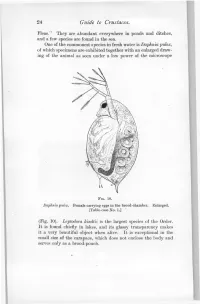
24 Guide to Crustacea
24 Guide to Crustacea. Fleas." They are abundant everywhere in ponds and ditches, and a few species are found in the sea. One of the commonest species in fresh water is Daphnia pulex, of which specimens are exhibited together with an enlarged draw- ing of the animal as seen under a low power of the microscope FIG. 10. Daphnia pulex. Female carrying eggs in the brood-chamber. Enlarged. [Table-case No. 1.] (Fig. 10). Leptodora kindtii is the largest species of the Order. It is found chiefly in lakes, and its glassy transparency makes it a very beautiful object when alive. It is exceptional in the small size of the carapace, which does not enclose the body and serves only as a brood-pouch. Ostracoda. 25 Sub-class II.—OSTRACODA. (Table-ease No. 1.) The number of somites, as indicated by the appendages, is smaller than in any other Crustacea, there being, at most, only two pairs of trunk-limbs behind those belonging to the head- region. The carapace forms a bivalved shell completely en- closing the body and limbs. There is a large, and often leg-like, palp on the mandible. The antennules and antennae are used for creeping or swimming. The Ostracoda (Fig. 10) are for the most part extremely minute animals, and only one or two of the larger species can be exhibited. They occur abundantly in fresh water and in FIG. 11. Shells of Ostracoda, seen from the side. A. Philomedes brenda (Myodocopa) ; B. Cypris fuscata (Podocopa); ('. Cythereis ornata (Podocopa): all much enlarged, n., Notch characteristic of the Myodocopa; e., the median eye ; a., mark of attachment of the muscle connecting the two valves of the shell. -

Epibiotic Associates of Oceanic-Stage Loggerhead Turtles from the Southeastern North Atlantic
Acknowledgements We thank the biology students of the occasional leatherback nests in Brazil. Marine Turtle Federal University of Paraíba (Pablo Riul, Robson G. dos Newsletter 96:13-16. Santos, André S. dos Santos, Ana C. G. P. Falcão, Stenphenson Abrantes, MS Elaine Elloy), the marathon MARCOVALDI, M. Â. & G. G. MARCOVALDI. 1999. Marine runner José A. Nóbrega, and the journalist Germana turtles of Brazil: the history and structure of Projeto Bronzeado for the volunteer field work; the Fauna department TAMAR-IBAMA. Biological Conservation 91:35-41. of IBAMA/PB and Jeremy and Diana Jeffers for kindly MARCOVALDI, M.Â., C.F. VIEITAS & M.H. GODFREY. 1999. providing photos, and also Alice Grossman for providing Nesting and conservation management of hawksbill turtles the TAMAR protocols. The manuscript benefited from the (Eretmochelys imbricata) in northern Bahia, Brazil. comments of two referees. Chelonian Conservation and Biology 3:301-307. BARATA, P.C.R. & F.F.C. FABIANO. 2002. Evidence for SAMPAIO, C.L.S. 1999. Dermochelys coriacea (Leatherback leatherback sea turtle (Dermochelys coriacea) nesting in sea turtle), accidental capture. Herpetological Review Arraial do Cabo, state of Rio de Janeiro, and a review of 30:38-39. Epibiotic Associates of Oceanic-Stage Loggerhead Turtles from the Southeastern North Atlantic Michael G. Frick1, Arnold Ross2, Kristina L. Williams1, Alan B. Bolten3, Karen A. Bjorndal3 & Helen R. Martins4 1 Caretta Research Project, P.O. Box 9841, Savannah, Georgia 31412 USA. (E-mail: [email protected]) 2 Scripps Institution of Oceanography, Marine Biology Research Division, La Jolla, California 92093-0202, USA, (E-mail: [email protected]) 3 Archie Carr Center for Sea Turtle Research and Department of Zoology, University of Florida, P.O. -

A Checklist of Turtle and Whale Barnacles
Journal of the Marine Biological Association of the United Kingdom, 2013, 93(1), 143–182. # Marine Biological Association of the United Kingdom, 2012 doi:10.1017/S0025315412000847 A checklist of turtle and whale barnacles (Cirripedia: Thoracica: Coronuloidea) ryota hayashi1,2 1International Coastal Research Center, Atmosphere and Ocean Research Institute, The University of Tokyo, 5-1-5, Kashiwanoha, Kashiwa-shi, Chiba 277-8564 Japan, 2Marine Biology and Ecology Research Program, Extremobiosphere Research Center, Japan Agency for Marine–Earth Science and Technology A checklist of published records of coronuloid barnacles (Cirripedia: Thoracica: Coronuloidea) attached to marine vertebrates is presented, with 44 species (including 15 fossil species) belonging to 14 genera (including 3 fossil genera) and 3 families recorded. Also included is information on their geographical distribution and the hosts with which they occur. Keywords: checklist, turtle barnacles, whale barnacles, Chelonibiidae, Emersoniidae, Coronulidae, Platylepadidae, host and distribution Submitted 10 May 2012; accepted 16 May 2012; first published online 10 August 2012 INTRODUCTION Superorder THORACICA Darwin, 1854 Order SESSILIA Lamarck, 1818 In this paper, a checklist of barnacles of the superfamily Suborder BALANOMORPHA Pilsbry, 1916 Coronuloidea occurring on marine animals is presented. Superfamily CORONULOIDEA Newman & Ross, 1976 The systematic arrangement used herein follows Newman Family CHELONIBIIDAE Pilsbry, 1916 (1996) rather than Ross & Frick (2011) for reasons taken up in Hayashi (2012) in some detail. The present author Genus Chelonibia Leach, 1817 deems the subfamilies of the Cheonibiidae (Chelonibiinae, Chelonibia caretta (Spengler, 1790) Emersoniinae and Protochelonibiinae) proposed by Harzhauser et al. (2011), as well as those included of Ross & Lepas caretta Spengler, 1790: 185, plate 6, figure 5. -

Cirripedia of Madeira
View metadata, citation and similar papers at core.ac.uk brought to you by CORE provided by Universidade do Algarve Helgol Mar Res (2006) 60: 207–212 DOI 10.1007/s10152-006-0036-5 ORIGINAL ARTICLE Peter Wirtz Æ Ricardo Arau´jo Æ Alan J. Southward Cirripedia of Madeira Received: 13 September 2005 / Revised: 12 January 2006 / Accepted: 13 January 2006 / Published online: 3 February 2006 Ó Springer-Verlag and AWI 2006 Abstract We give a list of Cirripedia from Madeira phers. The marine invertebrates have been less studied Island and nearby deep water, based on specimens in and there has been no compilation of cirripede records the collection of the Museu Municipal do Funchal for Madeira, comparable to those for the Azores (Histo´ria Natural) (MMF), records mentioned in the archipelago (Young 1998a; Southward 1999). We here literature, and recent collections. Tesseropora atlantica summarize records from Madeira and nearby deep water Newman and Ross, 1976 is recorded from Madeira for and discuss their biogeographical implications. the first time. The Megabalanus of Madeira is M. az- oricus. There are 20 genera containing 27 species, of which 22 occur in depths less than 200 m. Of these Methods shallow water species, eight are wide-ranging oceanic forms that attach to other organisms or to floating The records are based on (1) the work of R.T. Lowe, objects, leaving just 13 truly benthic shallow water who sent specimens to Charles Darwin; (2) material in barnacles. This low diversity is probably a consequence the Museu Municipal do Funchal (Histo´ria Natural) of the distance from the continental coasts and the (MMF); (3) casual collecting carried out by residents or small area of the available habitat. -

An Invitation to Monitor Georgia's Coastal Wetlands
An Invitation to Monitor Georgia’s Coastal Wetlands www.shellfish.uga.edu By Mary Sweeney-Reeves, Dr. Alan Power, & Ellie Covington First Printing 2003, Second Printing 2006, Copyright University of Georgia “This book was prepared by Mary Sweeney-Reeves, Dr. Alan Power, and Ellie Covington under an award from the Office of Ocean and Coastal Resource Management, National Oceanic and Atmospheric Administration. The statements, findings, conclusions, and recommendations are those of the authors and do not necessarily reflect the views of OCRM and NOAA.” 2 Acknowledgements Funding for the development of the Coastal Georgia Adopt-A-Wetland Program was provided by a NOAA Coastal Incentive Grant, awarded under the Georgia Department of Natural Resources Coastal Zone Management Program (UGA Grant # 27 31 RE 337130). The Coastal Georgia Adopt-A-Wetland Program owes much of its success to the support, experience, and contributions of the following individuals: Dr. Randal Walker, Marie Scoggins, Dodie Thompson, Edith Schmidt, John Crawford, Dr. Mare Timmons, Marcy Mitchell, Pete Schlein, Sue Finkle, Jenny Makosky, Natasha Wampler, Molly Russell, Rebecca Green, and Jeanette Henderson (University of Georgia Marine Extension Service); Courtney Power (Chatham County Savannah Metropolitan Planning Commission); Dr. Joe Richardson (Savannah State University); Dr. Chandra Franklin (Savannah State University); Dr. Dionne Hoskins (NOAA); Dr. Charles Belin (Armstrong Atlantic University); Dr. Merryl Alber (University of Georgia); (Dr. Mac Rawson (Georgia Sea Grant College Program); Harold Harbert, Kim Morris-Zarneke, and Michele Droszcz (Georgia Adopt-A-Stream); Dorset Hurley and Aimee Gaddis (Sapelo Island National Estuarine Research Reserve); Dr. Charra Sweeney-Reeves (All About Pets); Captain Judy Helmey (Miss Judy Charters); Jan Mackinnon and Jill Huntington (Georgia Department of Natural Resources). -

Checklist of the Australian Cirripedia
AUSTRALIAN MUSEUM SCIENTIFIC PUBLICATIONS Jones, D. S., J. T. Anderson and D. T. Anderson, 1990. Checklist of the Australian Cirripedia. Technical Reports of the Australian Museum 3: 1–38. [24 August 1990]. doi:10.3853/j.1031-8062.3.1990.76 ISSN 1031-8062 Published by the Australian Museum, Sydney naturenature cultureculture discover discover AustralianAustralian Museum Museum science science is is freely freely accessible accessible online online at at www.australianmuseum.net.au/publications/www.australianmuseum.net.au/publications/ 66 CollegeCollege Street,Street, SydneySydney NSWNSW 2010,2010, AustraliaAustralia ISSN 1031-8062 ISBN 0 7305 7fJ3S 7 Checklist of the Australian Cirripedia D.S. Jones. J.T. Anderson & D.l: Anderson Technical Reports of the AustTalfan Museum Number 3 Technical Reports of the Australian Museum (1990) No. 3 ISSN 1031-8062 Checklist of the Australian Cirripedia D.S. JONES', J.T. ANDERSON*& D.T. AND ER SON^ 'Department of Aquatic Invertebrates. Western Australian Museum, Francis Street. Perth. WA 6000, Australia 2School of Biological Sciences, University of Sydney, Sydney. NSW 2006, Australia ABSTRACT. The occurrence and distribution of thoracican and acrothoracican barnacles in Australian waters are listed for the first time since Darwin (1854). The list comprises 204 species. Depth data and museum collection data (for Australian museums) are given for each species. Geographical occurrence is also listed by area and depth (littoral, neuston, sublittoral or deep). Australian contributions to the biology of Australian cimpedes are summarised in an appendix. All listings are indexed by genus and species. JONES. D.S.. J.T. ANDERSON & D.T. ANDERSON,1990. Checklist of the Australian Cirripedia. -

Stalked Barnacles
*Manuscript Click here to view linked References Stalked barnacles (Cirripedia, Thoracica) from the Upper Jurassic (Tithonian) Kimmeridge Clay of Dorset, UK; palaeoecology and bearing on the evolution of living forms Andy Gale School of Earth and Environmental Sciences, University of Portsmouth, Burnaby Building, Burnaby Road, Portsmouth PO1 3QL; E-mail address: [email protected] A B S T R A C T New thoracican cirripede material from the Kimmeridge Clay (Upper Jurassic, Tithonian) is described. This includes a log, encrusted on the lower surface with hundreds of perfectly preserved, articulated specimens of Etcheslepas durotrigensis Gale, 2014, and fewer specimens of Concinnalepas costata (Withers, 1928). Some individuals are preserved in life position, hanging from the underside of the wood, and the material provides new morphological information on both species. It appears that Martillepas ovalis (Withers, 1928), which occurs at the same level (Freshwater Steps Stone Band, pectinatus Zone) attached preferentially to ammonites, whereas E. durotrigensis and C. costata used wood as a substrate for their epiplanktonic lifestyle. Two regurgitates containing abundant barnacle valves, mostly broken, and some bivalve fragments, have been found in the Kimmeridge Clay. These were produced by a fish grazing on epiplanktonic species, and are only the second example of regurgitates containing barnacle valves known from the fossil record. The evolution of modern barnacle groups is discussed in the light of the new Jurassic material as well as recently published molecular phylogenies. New clades defined herein are called the Phosphatothoracica, the Calamida and the Unilatera. Keywords Epiplanktonic barnacles Kimmeridge Clay predation 1. INTRODUCTION Amongst the most remarkable fossils collected by Steve Etches from the Kimmeridge Clay of Dorset are articulated stalked barnacles. -

UNIVERSITY of CALIFORNIA, SAN DIEGO Abundance and Ecological
UNIVERSITY OF CALIFORNIA, SAN DIEGO Abundance and ecological implications of microplastic debris in the North Pacific Subtropical Gyre A dissertation submitted in partial satisfaction of the requirements for the degree Doctor of Philosophy in Oceanography by Miriam Chanita Goldstein Committee in charge: Professor Mark D. Ohman, Chair Professor Lihini I. Aluwihare Professor Brian Goldfarb Professor Michael R. Landry Professor James J. Leichter 2012 Copyright Miriam Chanita Goldstein, 2012 All rights reserved. SIGNATURE PAGE The Dissertation of Miriam Chanita Goldstein is approved, and it is acceptable in quality and form for publication on microfilm and electronically: PAGE _____________________________________________________________________ _____________________________________________________________________ _____________________________________________________________________ _____________________________________________________________________ _____________________________________________________________________ Chair University of California, San Diego 2012 iii DEDICATION For my mother, who took me to the tidepools and didn’t mind my pet earthworms. iv TABLE OF CONTENTS SIGNATURE PAGE ................................................................................................... iii DEDICATION ............................................................................................................. iv TABLE OF CONTENTS ............................................................................................. v LIST OF FIGURES -

(II) : Cirripeds Found in the Vicinity of the Seto Marine Biological
Studies on Cirripedian Fauna of Japan (II) : Cirripeds Found in Title the Vicinity of the Seto Marine Biological Laboratory Author(s) Hiro, Fujio Memoirs of the College of Science, Kyoto Imperial University. Citation Ser. B (1937), 12(3): 385-478 Issue Date 1937-10-30 URL http://hdl.handle.net/2433/257864 Right Type Departmental Bulletin Paper Textversion publisher Kyoto University MEMorRs oF THE CoLLEGE oF SclENcE, KyoTo IM?ERIAL UNIvERSITy, SERIEs B, VoL. XII, No. 3, ART. 17, 1937 Studie$ on Cirripedian FauRa of Japan II. Cirripeds Found in the Vicinity ef the Seto Mavine Biological Laboxatory By Fajio HIRo (Seto Marine Biological Laboratory, Wakayarna-ken) With 43 Text-pt•gKres (Received April 21, l937) Introductien The purpose of the present paper is to describe the theracic cirripeds found in the waters around the Sete Marine Biological Laboratory. The material dealt with in this paper was collected almost entirely by myself during the period extending from the summer of 1930 up to the present time, except a few species ob- tained from the S6y6-maru Expedition undertaken by the Ircperial Fisheries Experimental Station during the years 1926-1930. Descrip- tions of the latter have already been given (HiRo, 1933a). The present material consists, with few exceptions, of specimens from the littoral zone and shallow ;vvater ; noRe of the specimens are irom deep water. However, I have paid special attention to the commensal forms from the ecological and fauRistic standpoint, and have thes been able to enumerate a comparatively large number of species in such a re- stricted area as this district. -

Illuminating Our World: an Essay on the Unraveling of the Species Problem, with Assistance from a Barnacle and a Goose
Humanities 2012, 1, 145–165; doi:10.3390/h1030145 OPEN ACCESS humanities ISSN 2076-0787 www.mdpi.com/journal/humanities Article Illuminating our World: An Essay on the Unraveling of the Species Problem, with Assistance from a Barnacle and a Goose John Buckeridge * and Rob Watts Earth & Oceanic Systems Group, RMIT University, Melbourne, GPO Box 2476, Australia * Author to whom correspondence should be addressed; E-Mail: [email protected]; Tel.: +61-399-252-009. Received: 18 July 2012; in revised form: 27 September 2012 / Accepted: 8 October 2012 / Published: 15 October 2012 Abstract: In order to plan for the future, we must understand the past. This paper investigates the manner in which both naturalists and the wider community view one of the most intriguing of all questions: what makes a species special? Consideration is given to the essentialist view—a rigid perspective and ancient, Aristotelian perspective—that all organisms are fixed in form and nature. In the middle of the 19th century, Charles Darwin changed this by showing that species are indeed mutable, even humans. Advances in genetics have reinforced the unbroken continuum between taxa, a feature long understood by palaeontologists; but irrespective of this, we have persisted in utilizing the ‗species concept‘—a mechanism employed primarily to understand and to manipulate the world around us. The vehicles used to illustrate this journey in perception are the barnacle goose (a bird), and the goose barnacle (a crustacean). The journey of these two has been entwined since antiquity—in folklore, religion, diet and even science. Keywords: species concept; organic evolution; history of biology; goose barnacles; barnacle geese; Aristotle; Charles Darwin; Linnaeus 1. -

Download PDF File
Scientific Note Record of coastal colonization of the Lepadid goose barnacle Lepas anatifera Linnaeus, 1758 (Crustacea: Cirripedia) at Arraial do Cabo, RJ 1 1,2 LUIS FELIPE SKINNER & DANIELLE FERNANDES BARBOZA 1Universidade do Estado do Rio de Janeiro (UERJ), Laboratório de Ecologia e Dinâmica Bêntica Marinha, São Gonçalo, Rio de Janeiro. Rua Francisco Portela 1470, Patronato, São Gonçalo, RJ. 24435-005. 2Bolsista PROATEC, UERJ. E-mail: [email protected] Abstract. In this note we record the occurence of Lepas anatifera (Cirripedia) on the hull of a coastal supply boat that operates only in restricted inshore waters of Arraial do Cabo. This species is usually found on tropical and subtropical oceanic waters and, at region, could be classified as transient species. Key words: marine fouling, barnacle, recruitment, inshore transport, species distribution, Cabo Frio upwelling Resumo. Registro de colonização costeira da craca pedunculada Lepas anatifera Linnaeus, 1758 (Crustacea: Cirripedia) na costa de Arraial do Cabo, RJ. Nesta nota nós registramos a ocorrência de Lepas anatifera (Cirripedia) no casco de uma pequena embarcação que opera exclusivamente em águas costeiras de Arraial do Cabo. Esta espécie é típica de águas oceânicas de regiões tropicais e subtropicais e pode ser classificada como transiente na região. Palavras chave: incrustação marinha, craca, recrutamento, transporte costeiro, distribuição de espécies, ressurgência de Cabo Frio Lepas anatifera Linnaeus, 1758 (Fig. 1) is a and not by larval preference. pedunculate goose barnacle with cosmopolitan At Cabo Frio region is frequent to find many distribution (Young 1990, 1998). They are found individuals that arrived to the coast on floating mainly on oceanic waters from tropical and debris.