Ida-Maria Jansson an Early Jurassic Conifer
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

6. Groundwater Resources
6. Groundwater Resources Contents 6 Groundwater Resources 6-1 6.1 Regulatory Framework 6-1 6.1.1 Queensland Legislation 6-1 6.2 Existing Environment 6-4 6.2.1 Project Location 6-4 6.2.2 Hydrology and Landforms 6-4 6.2.3 Geology 6-4 6.2.4 Hydrogeology 6-11 6.2.5 Groundwater Use 6-16 6.2.6 Existing Mine Groundwater Monitoring 6-25 6.2.7 Revised Project Baseline Groundwater Monitoring 6-27 6.2.8 Groundwater Levels 6-31 6.2.9 Groundwater Movement 6-35 6.2.10 Inter-aquifer Connectivity 6-38 6.2.11 Surface Water and Groundwater Interaction 6-39 6.2.12 Groundwater Quality 6-39 6.2.13 Environmental Values in the EPP Water 6-47 6.2.14 Conceptual Hydrogeological Model 6-49 6.3 Impact Assessment 6-53 6.3.1 Effects on Groundwater Levels 6-55 6.3.2 Effects on Groundwater Quality 6-72 6.3.3 Effects on Groundwater Users 6-75 6.3.4 Effects on Groundwater Dependent Ecosystems 6-77 6.4 Mitigation Measures 6-77 6.4.1 Groundwater Monitoring Program 6-77 6.4.2 Landholder Bores 6-83 6.4.3 Groundwater Impact Prediction, Validation and Review 6-83 6.4.4 Mitigation Measures for affected Groundwater Users 6-84 6.5 Conclusions 6-84 New Acland Coal Mine Stage 3 Project – Environmental Impact Statement PAGE i 6 Groundwater Resources This Chapter describes the groundwater resources that may be affected by the revised Project, how they might be affected, and the measures required for the mitigation of potential negative effects. -

The Mesozoic Megafossil Genus Linguifolium Arber 1917
Acta Palaeobotanica 55(2): 123–147, 2015 DOI: 10.1515/acpa-2015-0009 The Mesozoic megafossil genus Linguifolium Arber 1917 GARY A. PATTEMORE1, JOHN F. RIGBY 2 and GEOFFREY PLAYFORD 3 1 School of Earth Sciences, The University of Queensland, St. Lucia, Queensland 4072, Australia; e-mail: [email protected] 2 School of Earth, Environmental and Biological Sciences, Queensland University of Technology, GPO Box 2434, Brisbane, Queensland 4001, Australia; e-mail: [email protected] 3 School of Earth Sciences, The University of Queensland, St. Lucia, Queensland 4072, Australia; e-mail: [email protected] Received 24 April 2015; accepted for publication 3 September 2015 ABSTRACT. The plant megafossil genus Linguifolium Arber 1917 is chiefly known from the Middle and Upper Triassic of Gondwana. The range of Linguifolium extended beyond Gondwana by the Late Triassic, persisting there through the earliest Jurassic (Hettangian). The parent plants probably grew in a well-watered, canopied environment. Diagnoses of the genus and four of its species – Linguifolium tenison-woodsii (Shirley 1898) Retallack 1980, L. waitakiense Bell in Bell et al. 1956, L. parvum Holmes & Anderson in Holmes et al. 2010, and L. steinmannii (Solms-Laubach 1899) Arber 1917 – are emended with particular reference to venation and leaf morphology; consequently, the stratigraphic ranges of the species have been more precisely defined. Coalescent venation has previously been reported in some species of Linguifolium and is identified in new material described herein. Although the vast majority of specimens assigned to the genus are from the Upper Triassic, none shows coalescent venation. This character is entirely restricted to the Middle Triassic, in particu- lar to two species: L. -

Background Paper on New South Wales Geology with a Focus on Basins Containing Coal Seam Gas Resources
Background Paper on New South Wales Geology With a Focus on Basins Containing Coal Seam Gas Resources for Office of the NSW Chief Scientist and Engineer by Colin R. Ward and Bryce F.J. Kelly School of Biological, Earth and Environmental Sciences University of New South Wales Date of Issue: 28 August 2013 Our Reference: J083550 CONTENTS Page 1. AIMS OF THE BACKGROUND PAPER .............................................................. 1 1.1. SIGNIFICANCE OF AUSTRALIAN CSG RESOURCES AND PRODUCTION ................... 1 1.2. DISCLOSURE .................................................................................................... 2 2. GEOLOGY AND EVALUATION OF COAL AND COAL SEAM GAS RESOURCES ............................................................................................................. 3 2.1. NATURE AND ORIGIN OF COAL ........................................................................... 3 2.2. CHEMICAL AND PHYSICAL PROPERTIES OF COAL ................................................ 4 2.3. PETROGRAPHIC PROPERTIES OF COAL ............................................................... 4 2.4. GEOLOGICAL FEATURES OF COAL SEAMS .......................................................... 6 2.5. NATURE AND ORIGIN OF GAS IN COAL SEAMS .................................................... 8 2.6. GAS CONTENT DETERMINATION ........................................................................10 2.7. SORPTION ISOTHERMS AND GAS HOLDING CAPACITY .........................................11 2.8. METHANE SATURATION ....................................................................................12 -

Review of State Conservation Areas
Review of State Conservation Areas Report of the first five-year review of State Conservation Areas under the National Parks and Wildlife Act 1974 November 2008 Cover photos (clockwise from left): Trial Bay Goal, Arakoon SCA (DECC); Glenrock SCA (B. Peters, DECC); Banksia, Bent Basin SCA (M. Lauder, DECC); Glenrock SCA (B. Peters, DECC). © Copyright State of NSW and Department of Environment and Climate Change NSW. The Department of Environment and Climate Change NSW and State of NSW are pleased to allow this material to be reproduced for educational or non-commercial purposes in whole or in part, provided the meaning is unchanged and its source, publisher and authorship are acknowledged. Specific permission is required for the reproduction of photographs. Published by: Department of Environment and Climate Change 59–61 Goulburn Street PO Box A290 Sydney South 1232 Ph: (02) 9995 5000 (switchboard) Ph: 131 555 (environment information and publications requests) Ph: 1300 361 967 (national parks information and publications requests) Fax: (02) 9995 5999 TTY: (02) 9211 4723 Email: [email protected] Website: www.environment.nsw.gov.au ISBN 978-1-74122-981-3 DECC 2008/516 November 2008 Printed on recycled paper Contents Minister’s Foreword iii Part 1 – State Conservations Areas 1 State Conservation Areas 4 Exploration and mining in NSW 6 History and current trends 6 Titles 7 Assessments 7 Compliance and rehabilitation 8 Renewals 8 Exploration and mining in State Conservation Areas 9 The five-year review 10 Purpose of the review 10 -

PAPERS Department of Geology
PAPERS Department of Geology University of Queensland Volume 11 Number 4 PAPERS Department of Geology »University of Queensland VOLUME 11 NUMBER 4 The Tweed and Focal Peak Shield Volcanoes, Southeast Queensland and Northeast New South Wales . A. EWART, N.C. STEVENS and J.A. ROSS P. 1 - 82 1 THE TWEED AND FOCAL PEAK SHIELD VOLCANOES, SOUTHEAST QUEENSLAND AND NORTHEAST NEW SOUTH WALES by A. Ewart, N.C. Stevens and J.A, Ross ABSTRACT •Two overlapping shield volcanoes of Late Oligocène — Early Miocene age form mountainous country in southeast Queensland and northeast New South Wales. The basaltic-rhyolitic volcanic formations and the putonic rocks (gabbros, syenites, monzonites) of the central complexes are described with regard to field relations, mineralogy, geochem istry and petrogenesis. The Tweed Shield Volcano, centred on the plutonic complex of Mount Warning, comprises the Beechmont and Hobwee Basalts, their equivalents on the southern side (the Lismore and Blue Knob Basalts), and more localized rhyolite formations, the Binna Burra and Nimbin Rhyolites. The earlier Focal Peak Shield Volcano is preserved mainly on its eastern flanks, where the Albert Basalt and Mount Gillies Volcanics underlie the Beechmont Basalt. A widespread conglomerate formation separates formations of the two shield volcanoes. Mount Warning plutonic complex comprises various gabbros, syenite and monzonite with a syenite-trachyte-basalt ring-dyke, intrusive trachyandesite and comen dite dykes. The fine-grained granite of Mount Nullum and the basaltic sills of Mount Terragon are included with the complex. Each phase was fed by magma pulses from deeper chambers. Some degree of in situ crystal fractionation is shown by the gabbros, but the syenitic phase was already fractionated prior to emplacement. -

The Geology of NSW
The Geology of NSW The geological characteristics and history of NSW with a focus on coal seam gas (CSG) resources A report commissioned for the NSW Chief Scientist’s Office, May, 2013. Authors: Dr Craig O’Neill1, [email protected] Dr Cara Danis1, [email protected] 1Department of Earth and Planetary Science, Macquarie University, Sydney, NSW, 2109. Contents A brief glossary of terms i 1. Introduction 01 2. Scope 02 3. A brief history of NSW Geology 04 4. Evolution of the SydneyGunnedahBowen Basin System 16 5. Sydney Basin 19 6. Gunnedah Basin 31 7. Bowen Basin 40 8. Surat Basin 51 9. ClarenceMoreton Basin 60 10. Gloucester Basin 70 11. Murray Basin 77 12. Oaklands Basin 84 13. NSW Hydrogeology 92 14. Seismicity and stress in NSW 108 15. Summary and Synthesis 113 ii A brief Glossary of Terms The following constitutes a brief, but by no means comprehensive, compilation of some of the terms used in this review that may not be clear to a non‐geologist reader. Many others are explained within the text. Tectonothermal: The involvement of either (or both) tectonics (the large‐scale movement of the Earth’s crust and lithosphere), and geothermal activity (heating or cooling the crust). Orogenic: pertaining to an orogen, ie. a mountain belt. Associated with a collisional or mountain‐building event. Ma: Mega‐annum, or one million years. Conventionally associated with an age in geochronology (ie. million years before present). Epicratonic: “on the craton”, pertaining to being on a large, stable landmass (eg. -

Surat Underground Water Impact Report
Underground Water Impact Report for the Surat Cumulative Management Area 18 July 2012 Prepared by: Coal Seam Gas Water Queensland Water Commission © State of Queensland (Queensland Water Commission) 2012 Surat Underground Water Impact Report Contents Tables......................................................................................................................................... iv Figures........................................................................................................................................ iv Appendices..................................................................................................................................v Abbreviations.............................................................................................................................. vi Overview ................................................................................................................................ ix 1. Introduction ........................................................................................................................1 1.1 Water Rights.........................................................................................................................................1 1.2 Cumulative Management......................................................................................................................1 1.3 Surat Underground Water Impact Report .............................................................................................1 1.4 Implementation -

Malcolm Napier Thesis (PDF 10MB)
AN INTEGRATED HYDROLOGICAL AND HYDROCHEMICAL STUDY OF SURFACE AND GROUNDWATERS IN THE BUNGAWALBIN CREEK CATCHMENT, NORTHEAST NSW, AUSTRALIA. Malcolm Bruce Napier B.Nat.Res. School of Earth, Environmental and Biological Sciences Queensland University of Technology 2017 A thesis submitted for the degree of Master of Applied Science. Keywords Alluvium, Bungawalbin Creek, Bungawalbin Member, Clarence-Moreton Basin, coal-seam gas, Coraki Fault, down-hole geophysics, drill-stem test, fault hydrogeology, fracking, geophysics, Grafton Formation, groundwater, aquifer connectivity, groundwater dependent ecosystems, groundwater discharge, groundwater recharge, hydrochemistry, hydrology, stable isotope, Kangaroo Creek Sandstone, three-dimensional modelling, transient electromagnetics, Maclean Sandstone, methane, methane isotopes, Orara formation, petroleum, Piora Member, Rappville Member, tidal flow, Walloon Coal Measures i Abstract This study integrates hydrological, geophysical, geochemical and isotopic data to develop a conceptual model of surface and groundwater in the Bungawalbin Creek catchment, the southern sub-catchment of the Richmond River in north eastern New South Wales, Australia. The investigation brings together existing and newly-acquired data to provide new insights into the hydrogeology of the Clarence-Moreton Basin consolidated sediments as well as the South Casino Gravel shallow alluvial aquifer, and highlights connectivity between the aquifers and with surface water. Borehole stratigraphic data has been used to create a 3-D digital model of the consolidated sediments and has emphasized the presence of a major fault structure, the Coraki Fault. Hydrochemical and hydraulic evidence of recharge and discharge rates and localities in the alluvium is presented. These include potentiometric mapping, chloride mass-balance and major ion data, which indicate hydraulic continuity in the alluvial aquifer from the upper to the lower catchment. -

Early Cretaceous Macrofloras of Western Australia
Records of the Westem Australian Museum 18: 19-65 (1996), Early Cretaceous macrofloras of Western Australia Stephen McLoughlin School of Botany, The University of Melbourne, Parkville, Victoria 3052, Australia Abstract - Western Australian, Lower Cretaceous, macrofloras from the Broome Sandstone and Callawa Formation (Canning Basin), Nanutarra Formation and Birdrong Sandstone (Carnarvon Basin), Cronin Sandstone (Officer Basin), and Leederville and Bullsbrook Formations (Perth Basin) incorporate a range of lycophytes, ferns, pteridosperms, bennettitaleans, and conifers, The new monotypic genus Roebuckia is established for spatulate fern fronds (R, spatulata) of possible vittariacean alliance. Other newly established species include Phyllopteroides westralensis, Elatocladus ginginensis, and Carpolithes bullsbrookensis, Although the Western Australian fossil suites reveal some specific differences from other Australian late Mesozoic assemblages, several shared index taxa and the high proportion of bennettitaleans support correlation with the Victorian Neocomian Ptilophyllum-Pachypteris austropapillosa Zone (Zone B). Some Western Australian taxa are shared with Indian assemblages but fewer similarities exist with other Gondwanan Early Cretaceous floras, The representation of several hydrophilous fern, lycophyte, and pteridosperm groups together with growth indices from fossil woods implies a seasonal humid mesothermal climate for the Western Australian eratonic margin during the Neocomian-Barremian. Minor differences between the Western Australian -

Appendix D Surat Basin Geological Model Surat Basin Stratigraphic Framework April 2012
Appendix D Surat Basin geological model Surat Basin Stratigraphic Framework April 2012 Surat Basin Stratigraphic Framework April 2012 – Surat Basin Stratigraphic Framework April 2012 Table of Contents 1. Introduction .................................................................................................................................... 4 2. Methodology ................................................................................................................................... 8 2.1. Area of Interest ....................................................................................................................... 8 2.2. Datasets .................................................................................................................................. 8 2.3. Geological Modelling ............................................................................................................ 10 3. Regional Structure ........................................................................................................................ 12 4. Geological Descriptions ................................................................................................................. 17 4.1. Base Jurassic Unconformity .................................................................................................. 17 4.2. Precipice Sandstone .............................................................................................................. 17 4.3. Evergreen Formation ........................................................................................................... -

For Personal Use Only Use Personal For
ASX Announcement 30 April 2010 31 March 2010 - Quarterly Report Highlights • Kilgore agrees to acquire 51% of Clarence Moreton Resource Pty Ltd which has 2,050 km 2 of permits in the Clarence Moreton Basin of northern NSW that are prospective for coal seam gas and conventional hydrocarbons • Estimated potential gas in place in all permits of 7.3 TCF • Work commences on the Galveston 307 pipeline and production facilities and is expected to be completed in June 2010 Corporate Acquisition Kilgore entered into an agreement to acquire 51% of the issued capital of Clarence Moreton Resources Pty Ltd (CMR) that has 100% interest in three permits in the highly prospective Clarence Moreton Basin in New South Wales. CMR has 100% interest in both the conventional and coal seam rights of Petroleum Exploration Licenses (PEL) 457, 478 and 479 covering an area of approximately 2,050 km 2. The permit area is located close to planned gas infrastructure and adjacent to significant acreage positions held by major CSG/LNG companies. Subject to the satisfaction of certain terms and conditions, Kilgore proposes to acquire 51% of CMR through the issue of 175 million fully-paid ordinary Kilgore shares and 175 million options to acquire Kilgore shares at $0.05 on or before 30 th June 2012. Kilgore has also secured an option to acquire the remaining 49% of CMR through the issue of additional shares and options and the payment of an annual royalty on gross sales arising from the project. Coal Seam Gas Potential The project is well located with recent coal seam gas exploration success in the Casino area. -
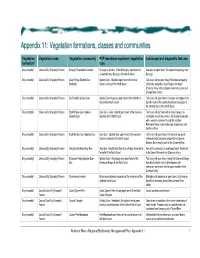
Vegetation Formations, Classes and Communities
Appendix 11: Vegetatiion formatiions, cllasses and communiities Vegetation Vegetation class Vegetation community PVP developer equivalent vegetation Landscape and diagnostic features formation* type Dry sclerophyll Clarence Dry Sclerophyll Forests Baryulgil Serpentinite Complex Eucalyptus ophitica - White Mahogany open forest on Low open or open forest. On serpentinite geology near serpentinite near Baryulgil of the North Coast Baryulgil. Dry sclerophyll Clarence Dry Sclerophyll Forests Coast Range Spotted Gum- Spotted Gum - Blackbutt open forest of the lower Tall to very tall dry open forest. Restricted and patchy Blackbutt Clarence Valley of the North Coast distribution along the Coast Range in the lower Clarence Valley, with a disjunct western occurrence in Grange State Forest. Dry sclerophyll Clarence Dry Sclerophyll Forests Dry Foothills Spotted Gum Spotted Gum dry grassy open forest of the foothills of Tall to very tall open forest. On slopes and ridges of dry the northern North Coast foothills areas of the coastal hinterland and gorges of the northern parts of the North Coast. Dry sclerophyll Clarence Dry Sclerophyll Forests Foothill Grey Gum-Ironbark- Grey Gum - Grey Ironbark open forest of the Clarence Tall to very tall dry forest with a mixed canopy. On Spotted Gum lowlands of the North Coast sandstone and siliceous soils in the Clarence lowlands with a western extension through the southern Richmond Range inland to Ewingar State Forest and the Mann River. Dry sclerophyll Clarence Dry Sclerophyll Forests Foothills Grey Gum-Spotted Gum Grey Gum - Spotted Gum open forest of the southern Tall to very tall open forest. On high and low quartz Clarence lowlands of the North Coast sediments in the southern portion of the Clarence- Moreton Basin mainly south of the Clarence River.