104612 Sletvoldtrunschkewimm
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

1.2 RP TVB ANNEXE ABT Especes Et Enjeux
La connaissance de la biodiversité sur l’Est Cantal un enjeu pour le développement du territoire ! Depuis avril 2017, le SYTEC s’est engagé dans une démarche d’Atlas de la Biodiversité Territoriale, avec l’objectif d’une part, de mieux connaitre la biodiversité du territoire et, d’autre part, d’intégrer les enjeux connus dans les démarches de planification notamment. Dans ce cadre, un ensemble de données naturalistes sur les territoires de l’Est Cantal ont pu être collectées auprès de nombreuses structures (Muséum National d’Histoire Naturelle, collectivités, associations, naturalistes professionnels…). Des inventaires sont également confiés chaque année par le SYTEC à des structures professionnelles pour compléter les connaissances sur des secteurs mal connus. En complément, des enquêtes participatives, qui rencontrent un vif succès, ont été lancées à destination du grand public, particulièrement des habitants des territoires. Ce travail de collecte a permis de réunir plus d’un million de données depuis le début de la démarche. A la fin de l’année 2018, ce sont 2505 espèces qui ont été répertoriés sur le territoire depuis 10 ans pour la faune (1017 espèces) et au cours de ces 15 dernières années pour la flore, les champignons et les algues (respectivement 1359, 31 et 98 espèces recensés). Parmi elles, 303 espèces sont considérées comme des espèces à enjeux en termes de préservation. Les inventaires se sont poursuivis en 2019 et continueront en 2020, permettant une mise à jour annuelle des connaissances de l’Est Cantal. Afin de partager -

Check List of Noctuid Moths (Lepidoptera: Noctuidae And
Бiологiчний вiсник МДПУ імені Богдана Хмельницького 6 (2), стор. 87–97, 2016 Biological Bulletin of Bogdan Chmelnitskiy Melitopol State Pedagogical University, 6 (2), pp. 87–97, 2016 ARTICLE UDC 595.786 CHECK LIST OF NOCTUID MOTHS (LEPIDOPTERA: NOCTUIDAE AND EREBIDAE EXCLUDING LYMANTRIINAE AND ARCTIINAE) FROM THE SAUR MOUNTAINS (EAST KAZAKHSTAN AND NORTH-EAST CHINA) A.V. Volynkin1, 2, S.V. Titov3, M. Černila4 1 Altai State University, South Siberian Botanical Garden, Lenina pr. 61, Barnaul, 656049, Russia. E-mail: [email protected] 2 Tomsk State University, Laboratory of Biodiversity and Ecology, Lenina pr. 36, 634050, Tomsk, Russia 3 The Research Centre for Environmental ‘Monitoring’, S. Toraighyrov Pavlodar State University, Lomova str. 64, KZ-140008, Pavlodar, Kazakhstan. E-mail: [email protected] 4 The Slovenian Museum of Natural History, Prešernova 20, SI-1001, Ljubljana, Slovenia. E-mail: [email protected] The paper contains data on the fauna of the Lepidoptera families Erebidae (excluding subfamilies Lymantriinae and Arctiinae) and Noctuidae of the Saur Mountains (East Kazakhstan). The check list includes 216 species. The map of collecting localities is presented. Key words: Lepidoptera, Noctuidae, Erebidae, Asia, Kazakhstan, Saur, fauna. INTRODUCTION The fauna of noctuoid moths (the families Erebidae and Noctuidae) of Kazakhstan is still poorly studied. Only the fauna of West Kazakhstan has been studied satisfactorily (Gorbunov 2011). On the faunas of other parts of the country, only fragmentary data are published (Lederer, 1853; 1855; Aibasov & Zhdanko 1982; Hacker & Peks 1990; Lehmann et al. 1998; Benedek & Bálint 2009; 2013; Korb 2013). In contrast to the West Kazakhstan, the fauna of noctuid moths of East Kazakhstan was studied inadequately. -

Diversity of the Moth Fauna (Lepidoptera: Heterocera) of a Wetland Forest: a Case Study from Motovun Forest, Istria, Croatia
PERIODICUM BIOLOGORUM UDC 57:61 VOL. 117, No 3, 399–414, 2015 CODEN PDBIAD DOI: 10.18054/pb.2015.117.3.2945 ISSN 0031-5362 original research article Diversity of the moth fauna (Lepidoptera: Heterocera) of a wetland forest: A case study from Motovun forest, Istria, Croatia Abstract TONI KOREN1 KAJA VUKOTIĆ2 Background and Purpose: The Motovun forest located in the Mirna MITJA ČRNE3 river valley, central Istria, Croatia is one of the last lowland floodplain 1 Croatian Herpetological Society – Hyla, forests remaining in the Mediterranean area. Lipovac I. n. 7, 10000 Zagreb Materials and Methods: Between 2011 and 2014 lepidopterological 2 Biodiva – Conservation Biologist Society, research was carried out on 14 sampling sites in the area of Motovun forest. Kettejeva 1, 6000 Koper, Slovenia The moth fauna was surveyed using standard light traps tents. 3 Biodiva – Conservation Biologist Society, Results and Conclusions: Altogether 403 moth species were recorded Kettejeva 1, 6000 Koper, Slovenia in the area, of which 65 can be considered at least partially hygrophilous. These results list the Motovun forest as one of the best surveyed regions in Correspondence: Toni Koren Croatia in respect of the moth fauna. The current study is the first of its kind [email protected] for the area and an important contribution to the knowledge of moth fauna of the Istria region, and also for Croatia in general. Key words: floodplain forest, wetland moth species INTRODUCTION uring the past 150 years, over 300 papers concerning the moths Dand butterflies of Croatia have been published (e.g. 1, 2, 3, 4, 5, 6, 7, 8). -

Database of Irish Lepidoptera. 1 - Macrohabitats, Microsites and Traits of Noctuidae and Butterflies
Database of Irish Lepidoptera. 1 - Macrohabitats, microsites and traits of Noctuidae and butterflies Irish Wildlife Manuals No. 35 Database of Irish Lepidoptera. 1 - Macrohabitats, microsites and traits of Noctuidae and butterflies Ken G.M. Bond and Tom Gittings Department of Zoology, Ecology and Plant Science University College Cork Citation: Bond, K.G.M. and Gittings, T. (2008) Database of Irish Lepidoptera. 1 - Macrohabitats, microsites and traits of Noctuidae and butterflies. Irish Wildlife Manual s, No. 35. National Parks and Wildlife Service, Department of the Environment, Heritage and Local Government, Dublin, Ireland. Cover photo: Merveille du Jour ( Dichonia aprilina ) © Veronica French Irish Wildlife Manuals Series Editors: F. Marnell & N. Kingston © National Parks and Wildlife Service 2008 ISSN 1393 – 6670 Database of Irish Lepidoptera ____________________________ CONTENTS CONTENTS ........................................................................................................................................................1 ACKNOWLEDGEMENTS ....................................................................................................................................1 INTRODUCTION ................................................................................................................................................2 The concept of the database.....................................................................................................................2 The structure of the database...................................................................................................................2 -

The Lepidoptera of Formby
THE RAVEN ENTOMOLOGICAL — AND — NATURAL HISTORY SOCIETY FOUNDED 1946 THE LEPIDOPTERA OF FORMBY Price: TWO SHILLINGS & SIXPENCE THE RAVEN ENTOMOLOGICAL AND NATURAL HISTORY SOCIETY FOUNDED 1946 THE LEPIDOPTERA OF FORMBY — B Y — M. J. LEECH H. N. MICHAELIS With a short biographical note on the late G. de C. Fraser by C. de C. Whiteley For us the wide open spaces, the mountams and valleys, the old walls and the hedges and ditches, wherein lie adventure and interest for to-day, to-morrow, and a lifetime. n Printed by T. B unci-e & Co. L td., Arbroath. GKRALI) i)E C. FRASER rOHEWORl) FOREWORD BY AI,LAN BRJNDLK TT was in August, 1939, that T first liad the pleasure of meeting the Frasers. Together with a small party of entomologists from N.E. I.ancashire. invited to eollect at light near the shore at Formby, I experienced the somewhat overwhelming enthrisiasm and hospitality extended to all at “ Warren Mount” . Fed, feted, and equipped, we were taken by cars to the shore, sheets were laid down in front of the headlights, and a memorable night ensued. The night was dark and warm, the moths arrived in great numbers and, true to the Fraser tradition, work did not cease until a few minutes before the last train left Formby, when a hurried dash to the station deposited a happy party of entomologists on the first stage of the journey home. The next meeting was long delayed. The following week-end saw the black-out in force, and it was not until 1946 that T found the Frasens, still enthusiastic, establishing the Eaven Society. -

Scottish Macro-Moth List, 2015
Notes on the Scottish Macro-moth List, 2015 This list aims to include every species of macro-moth reliably recorded in Scotland, with an assessment of its Scottish status, as guidance for observers contributing to the National Moth Recording Scheme (NMRS). It updates and amends the previous lists of 2009, 2011, 2012 & 2014. The requirement for inclusion on this checklist is a minimum of one record that is beyond reasonable doubt. Plausible but unproven species are relegated to an appendix, awaiting confirmation or further records. Unlikely species and known errors are omitted altogether, even if published records exist. Note that inclusion in the Scottish Invertebrate Records Index (SIRI) does not imply credibility. At one time or another, virtually every macro-moth on the British list has been reported from Scotland. Many of these claims are almost certainly misidentifications or other errors, including name confusion. However, because the County Moth Recorder (CMR) has the final say, dubious Scottish records for some unlikely species appear in the NMRS dataset. A modern complication involves the unwitting transportation of moths inside the traps of visiting lepidopterists. Then on the first night of their stay they record a species never seen before or afterwards by the local observers. Various such instances are known or suspected, including three for my own vice-county of Banffshire. Surprising species found in visitors’ traps the first time they are used here should always be regarded with caution. Clerical slips – the wrong scientific name scribbled in a notebook – have long caused confusion. An even greater modern problem involves errors when computerising the data. -

Tarset and Greystead Biological Records
Tarset and Greystead Biological Records published by the Tarset Archive Group 2015 Foreword Tarset Archive Group is delighted to be able to present this consolidation of biological records held, for easy reference by anyone interested in our part of Northumberland. It is a parallel publication to the Archaeological and Historical Sites Atlas we first published in 2006, and the more recent Gazeteer which both augments the Atlas and catalogues each site in greater detail. Both sets of data are also being mapped onto GIS. We would like to thank everyone who has helped with and supported this project - in particular Neville Geddes, Planning and Environment manager, North England Forestry Commission, for his invaluable advice and generous guidance with the GIS mapping, as well as for giving us information about the archaeological sites in the forested areas for our Atlas revisions; Northumberland National Park and Tarset 2050 CIC for their all-important funding support, and of course Bill Burlton, who after years of sharing his expertise on our wildflower and tree projects and validating our work, agreed to take this commission and pull everything together, obtaining the use of ERIC’s data from which to select the records relevant to Tarset and Greystead. Even as we write we are aware that new records are being collected and sites confirmed, and that it is in the nature of these publications that they are out of date by the time you read them. But there is also value in taking snapshots of what is known at a particular point in time, without which we have no way of measuring change or recognising the hugely rich biodiversity of where we are fortunate enough to live. -

Journal of Avian Biology JAV-01128 Valtonen, A., Latja, R., Leinonen, R
Journal of Avian Biology JAV-01128 Valtonen, A., Latja, R., Leinonen, R. and Pöysä, H. 2016. Arrival and onset of breeding of three passerine birds in eastern Finland tracks climatic variation and phenology of insects. – J. Avian Biol. doi: 10.1111/ jav.01128 Supplementary material 1 2 3 4 Supplementary material 5 6 Appendix 1. Are first arrival or onset of breeding related to population density? 7 We explored the possibility that the first arrival or onset of breeding are related to population 8 density (e.g., Dunn and Winkler 1999, Tryjanowski and Sparks 2001, Miller-Rushing et al. 2008, 9 Lindén 2011), which might confound the results found. Over the past decades, the breeding 10 populations of the Pied Flycatcher (50% increase in 38 years) and the Great Tit (population doubled 11 in 37 years) and the northern populations of the Common Redstart (population doubled in 35 years) 12 in Finland have significantly increased, while no long-term trend has been found of the Southern 13 populations of the Common Redstart (see population trajectories in Väisänen and Lehikoinen 14 2013). To address these questions, we added the annual breeding population size index (Väisänen 15 and Lehikoinen 2013) of each species as the fifth explanatory variable in the multiple regressions. 16 Nationwide estimates for the Pied Flycatcher and the Great Tit were used, but for the Common 17 Redstart, the population trajectories differed between Southern and Northern Finland; we therefore 18 used the population trajectory of Southern Finland in our analyses. These models were fitted for 19 data of years for which population index data was available (1975-2010 for the Pied Flycatcher, 20 1979-2010 for the Common Redstart, and 1976-2010 for the Great Tit). -

What Moths Fly in Winter? the Assemblage of Moths Active in a Temperate Deciduous Forest During the Cold Season in Central Poland
J. Entomol. Res. Soc., 17(2): 59-71, 2015 ISSN:1302-0250 What Moths Fly in Winter? The Assemblage of Moths Active in a Temperate Deciduous Forest During the Cold Season in Central Poland Jacek HIKISZ1 Agnieszka SOSZYŃSKA-MAJ2* Department of Invertebrate Zoology and Hydrobiology, University of Lodz, Banacha 12/16, 90-237 Łódź, POLAND, e-mails: 1 [email protected], 2*[email protected] ABSTRACT The composition and seasonal dynamics of the moth assemblage active in a temperate deciduous forest of Central Poland in autumn and spring was studied in two seasons 2007/2008 and 2008/2009. The standard light trapping method was used and, in addition, tree trunks were searched for resting moths. 42 species of moths from six families were found using both methods. The family Geometridae was predominant in terms of the numbers of individuals collected. Two geometrid species - Alsophila aescularia and Operophtera brumata - were defined as characteristic of the assemblage investigated. Late autumn and spring were richest in the numbers of species, whereas the species diversity was the lowest in mid-winter. Regression analysis showed that a temperature rise increased the species diversity of Geometridae but that rising air pressure negatively affected the abundance of Noctuidae. Key words: Lepidoptera, Geometridae, Noctuidae, autumn-spring activity, winter, phenology, atmospheric conditions, regression, Central Poland. INTRODUCTION The seasonal weather changes in a temperate climate have a great impact on poikilothermic animals, as they have a limited ability to regulate their body temperature. Thus, the earliest and most easily detectable response to climate change is an adjustment of species phenology (Huntley, 2007). -

Noctuoid Moths (Lepidoptera, Noctuoidea) of the Southern Part of Karakalpakstan
SJIF Impact Factor: 6.260| ISI I.F.Value:1.241| Journal DOI: 10.36713/epra2016 ISSN: 2455-7838(Online) EPRA International Journal of Research and Development (IJRD) Volume: 5 | Issue: 4 | April 2020 - Peer Reviewed Journal NOCTUOID MOTHS (LEPIDOPTERA, NOCTUOIDEA) OF THE SOUTHERN PART OF KARAKALPAKSTAN Bekchanov Khudaybergan Urinovich Docent, Faculty of Pedagogy, Department of methodology of pre-school education, Urgench State University Bekchanova Mohira Khudaybergan qizi PhD student of Khorezm Ma’mun Academy ABSTRACT The paper presents the results of faunal studies of the Lepidoptera group Noctuoidea in the southern part of Karakalpakstan, as well as a review of the literature on this topic. An annotated list of 51 species of 3 families is given: Erebidae, Nolidae and Noctuidae, which also includes previously published data on finds in the region. KEY WORDS: noctuoid, moths, Lepidoptera, Erebidae, Nolidae, Noctuidae, Karakalpakstan, ephemera, ephemerids, bush INTRODUCTION Sviridov, 2003; Kononenko, 2005; Matov et al., 2008; The family of Noctuidae is the largest family of Kononenko, 2010). This work is devoted to the study of Lepidoptera. About 30 thousand species are known in the the fauna of Lepidaptera in the southern part of world fauna, and this figure can hardly be considered Karakalpakstan in the spring-summer period. final. Uzbekistan stands out for its significant species abundance and zoogeographic diversity of the moths. The AREA OF THE STUDY study of the Noctuidae fauna of Uzbekistan has always The studies were conducted in the southern part been associated with the study of the Lepidoptera fauna of Karakalpakstan. Ephemeral plants and ephemeroids, of the Khorezm oasis. -

Vodič Za Preliminarnu Determinaciju Gusenica U Srbiji Na Osnovu Morfoloških, Bihejvioralnih I Ekoloških Karaktera
UNIVERZITET U NIŠU PRIRODNO-MATEMATIČKI FAKULTET NIŠ DEPARTMAN ZA BIOLOGIJU I EKOLOGIJU Master rad Vodič za preliminarnu determinaciju gusenica u Srbiji na osnovu morfoloških, bihejvioralnih i ekoloških karaktera Kandidat: Mentor: Aleksandra Trajković 297 prof. dr Vladimir Žikić Niš, 2020. UNIVERSITY OF NIŠ FACULTY OF SCIENCE AND MATHEMATICS DEPARTMENT OF BIOLOGY AND ECOLOGY Master thesis Preliminary Identification Guide to Caterpillars of Serbia Based on Morphological, Behavioral and Ecological Traits Candidate: Mentor: Aleksandra Trajković 297 Prof. Vladimir Žikić, PhD Niš, 2020. Dr Vladimiru Žikiću: Profesore, hvala Vam što ste mi mentor. Sub- i superfamiliji, kolegama, prijateljima i svima kojima su zbog mene gusenice postale tema razgovora, objekat (blagog) interesovanja i uopšte neizostavan deo svakodnevice (prevashodno enterijera): Hvala vam što se kladite na mene. Biografija Aleksandra Trajković rođena je u Nišu, 9. novembra 1996. godine. Nakon završene osnovne škole, upisuje medicinsku školu dr Milenko Hadžić u Nišu, smer fizioterapeutski tehničar, i završava je sa odličnim uspehom. Osnovne akademske studije upisuje 2015. godine na Prirodno-matematičkom fakultetu u Nišu, na Departmanu za biologiju i ekologiju, koje završava u junu 2018. godine prosekom 9,00. Iste godine, upisuje master akademske studije na Departmanu za biologiju i ekologiju, smer Biologija. Letnji semestar akademske 2018/19 provodi na Erasmus+ programu studentske mobilnosti, na Univerzitetu u Bjalistoku, Poljska, na Institutu za biologiju i hemiju, koji završava sa najvišim ocenama. Master akademske studije završava u matičnoj ustanovi, prosečnom ocenom 9,94. Sažetak Identifikacija gusenica do nivoa pripadnosti užoj taksonomskoj kategoriji otežana je diverzitetom taksona u okviru reda Lepidoptera, kao i nedostatkom odgovarajuće literature za determinaciju. Heterogenost u morfologiji i ekologiji ove grupe organizama ukazuju na izuzetan biološki značaj, ali i na kompleksnost precizne identifikacije. -
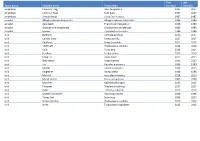
Taxon Group Common Name Taxon Name First Recorded Last
First Last Taxon group Common name Taxon name recorded recorded amphibian Common Frog Rana temporaria 1987 2017 amphibian Common Toad Bufo bufo 1987 2017 amphibian Smooth Newt Lissotriton vulgaris 1987 1987 annelid Alboglossiphonia heteroclita Alboglossiphonia heteroclita 1986 1986 annelid duck leech Theromyzon tessulatum 1986 1986 annelid Glossiphonia complanata Glossiphonia complanata 1986 1986 annelid leeches Erpobdella octoculata 1986 1986 bird Bullfinch Pyrrhula pyrrhula 2016 2017 bird Carrion Crow Corvus corone 2017 2017 bird Chaffinch Fringilla coelebs 2015 2017 bird Chiffchaff Phylloscopus collybita 2014 2016 bird Coot Fulica atra 2014 2014 bird Fieldfare Turdus pilaris 2015 2015 bird Great Tit Parus major 2015 2015 bird Grey Heron Ardea cinerea 2013 2017 bird Jay Garrulus glandarius 1999 1999 bird Kestrel Falco tinnunculus 1999 2015 bird Kingfisher Alcedo atthis 1986 1986 bird Mallard Anas platyrhynchos 2014 2015 bird Marsh Harrier Circus aeruginosus 2000 2000 bird Moorhen Gallinula chloropus 2015 2015 bird Pheasant Phasianus colchicus 2017 2017 bird Robin Erithacus rubecula 2017 2017 bird Spotted Flycatcher Muscicapa striata 1986 1986 bird Tawny Owl Strix aluco 2006 2015 bird Willow Warbler Phylloscopus trochilus 2015 2015 bird Wren Troglodytes troglodytes 2015 2015 bird Yellowhammer Emberiza citrinella 2000 2000 conifer Douglas Fir Pseudotsuga menziesii 2004 2004 conifer European Larch Larix decidua 2004 2004 conifer Lawson's Cypress Chamaecyparis lawsoniana 2004 2004 conifer Scots Pine Pinus sylvestris 1986 2004 crustacean