This PDF File Is Subject to the Following Conditions and Restrictions
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Impact Structures and Events – a Nordic Perspective
107 by Henning Dypvik1, Jüri Plado2, Claus Heinberg3, Eckart Håkansson4, Lauri J. Pesonen5, Birger Schmitz6, and Selen Raiskila5 Impact structures and events – a Nordic perspective 1 Department of Geosciences, University of Oslo, P.O. Box 1047, Blindern, NO 0316 Oslo, Norway. E-mail: [email protected] 2 Department of Geology, University of Tartu, Vanemuise 46, 51014 Tartu, Estonia. 3 Department of Environmental, Social and Spatial Change, Roskilde University, P.O. Box 260, DK-4000 Roskilde, Denmark. 4 Department of Geography and Geology, University of Copenhagen, Øster Voldgade 10, DK-1350 Copenhagen, Denmark. 5 Division of Geophysics, University of Helsinki, P.O. Box 64, FIN-00014 Helsinki, Finland. 6 Department of Geology, University of Lund, Sölvegatan 12, SE-22362 Lund, Sweden. Impact cratering is one of the fundamental processes in are the main reason that the Nordic countries are generally well- the formation of the Earth and our planetary system, as mapped. reflected, for example in the surfaces of Mars and the Impact craters came into the focus about 20 years ago and the interest among the Nordic communities has increased during recent Moon. The Earth has been covered by a comparable years. The small Kaalijärv structure of Estonia was the first impact number of impact scars, but due to active geological structure to be confirmed in northern Europe (Table 1; Figures 1 and processes, weathering, sea floor spreading etc, the num- 7). First described in 1794 (Rauch), the meteorite origin of the crater ber of preserved and recognized impact craters on the field (presently 9 craters) was proposed much later in 1919 (Kalju- Earth are limited. -

Impact Melt Rocks in the "Cretaceous
Large Meteorite Impacts (2003) alpha_t-z.pdf Contents — T through Z Locations and Compositions of Mare Ponds in South Pole-Aitken Basin on the Moon and Its Implication to the Impact Tectonics T. Takata and S. Hori ............................................................................................................................. 4058 New Laboratory Results on Field Sections at the Impact Crater of Araguainha (MT, GO, Brazil). Area of Proximal and Distal Impact Ejecta, Including Microspherules Dated from the End of Permian J. M. Théry, A. Crosta, E. Veto Akos, E. Bilal, K. Gal-Solymos, and E. Dransart................................. 4096 Techniques of Shock Wave Experiments and Determination of Hugoniot Data of Solids K. Thoma................................................................................................................................................ 4134 Differential Stress-controlled Deformation of Quartz During and After Hypervelocity Impact — Microstructural Evidence from the Charlevoix Impact Structure, Quebéc, Canada C. A. Trepmann and J. G. Spray............................................................................................................. 4026 Mjølnir Marine Crater Resulting from Oblique Impact: Compelling Evidence F. Tsikalas and J. I. Faleide................................................................................................................... 4005 A New Mid- to Late-Maastrichtian Impact in the Raton Basin 100m Below the K/T Boundary P. Turner, S. C. Sherlock, P. Clarke, and -

Polygonal Impact Craters in Argyre Region, Mars: Implications for Geology and Cratering Mechanics
Meteoritics & Planetary Science 43, Nr 10, 1605–1628 (2008) Abstract available online at http://meteoritics.org Polygonal impact craters in Argyre region, Mars: Implications for geology and cratering mechanics T. ÖHMAN1, 2*, M. AITTOLA2, V.-P. KOSTAMA2, J. RAITALA2, and J. KORTENIEMI2, 3 1Department of Geosciences, Division of Geology, University of Oulu, P.O. Box 3000, FI-90014, Finland 2Department of Physical Sciences, Division of Astronomy, University of Oulu, P.O. Box 3000, FI-90014, Finland 3Institut für Planetologie, Westfälische Wilhelms-Universität, Wilhelm-Klemm-Strasse 10, D-48149 Münster, Germany *Corresponding author. E-mail: [email protected] (Received 20 November 2007; revision accepted 15 April 2008) Abstract–Impact craters are not always circular; sometimes their rims are composed of several straight segments. Such polygonal impact craters (PICs) are controlled by pre-existing target structures, mainly faults or other similar planes of weakness. In the Argyre region, Mars, PICs comprise ∼17% of the total impact crater population (>7 km in diameter), and PICs are relatively more common in older geologic units. Their formation is mainly controlled by radial fractures induced by the Argyre and Ladon impact basins, and to a lesser extent by the basin-concentric fractures. Also basin-induced conjugate shear fractures may play a role. Unlike the PICs, ridges and graben in the Argyre region are mostly controlled by Tharsis-induced tectonism, with the ridges being concentric and graben radial to Tharsis. Therefore, the PICs primarily reflect an old impact basin-centered tectonic pattern, whereas Tharsis-centered tectonism responsible for the graben and the ridges has only minor influence on the PIC rim orientations. -

Chapter 6 ______Chapter 6
Chapter 6 _____________________________________________________________________ Chapter 6 Discussion 6.1 Introduction Previous structural studies of the collar rocks of the Vredefort Dome focused predominantly on specific areas of the collar (e.g., Lilly 1978, 1981) or specific features, such as pseudotachylitic breccias (e.g., Martini 1991; Gibson et al. 1997) and shatter cones (e.g., Manton 1962; Albat and Mayer 1989; Nicolaysen and Reimold 1999). Many of the studies were carried out in the era leading up to the confirmation of the dome as part of an impact structure and, hence, they did not necessarily examine these structures within the context of an impact-induced evolutionary history. This study aimed to document all structural features over the entire exposed extent of the collar of the dome in an attempt to establish geometric and/or temporal relationships between individual structures and their relation to the impact process (impact-induced versus non-impact-induced deformation). Furthermore, the temporal relationships between impact-related deformation features were investigated in order to obtain a better understanding of the processes involved in the evolution of the deep central parts of a complex impact structure during the cratering process. More specifically, this study aimed at improving the understanding of the processes that take place during the formation and subsequent collapse of the central uplift in such a crater. A brief summary of the main observations will be provided at the beginning of each section in order to point out the salient facts, which are necessary for a comprehensive discussion of the results with regard to the formation and collapse of the Vredefort Dome, as well as other impact structures in general. -

Giant Comets and Mass Extinctions of Life
Mon. Not. R. Astron. Soc. 000, ?? – ?? (2015) Printed 17 March 2015 (MN LATEX style file v2.2) Giant comets and mass extinctions of life W.M. Napier1⋆ 1Buckingham Centre for Astrobiology, University of Buckingham, Buckingham MK18 1EG, UK Accepted 2014 December 16. Received 2014 December 12; in original form 2014 March 31 ABSTRACT I find evidence for clustering in age of well-dated impact craters over the last 500 Myr. At least nine impact episodes are identified, with durations whose upper limits are set by the dating accuracy of the craters. Their amplitudes and frequency are inconsis- tent with an origin in asteroid breakups or Oort cloud disturbances, but are consistent with the arrival and disintegration in near-Earth orbits of rare, giant comets, mainly in transit from the Centaur population into the Jupiter family and Encke regions. About 1 in 10 Centaurs in Chiron-like orbits enter Earth-crossing epochs, usually repeatedly, each such epoch being generally of a few thousand years duration. On time-scales of geological interest, debris from their breakup may increase the mass of the near-Earth interplanetary environment by two or three orders of magnitude, yielding repeated episodes of bombardment and stratospheric dusting. I find a strong correlation between these bombardment episodes and major biostratigraphic and geo- logical boundaries, and propose that episodes of extinction are most effectively driven by prolonged encounters with meteoroid streams during bombardment episodes. Pos- sible mechanisms are discussed. Key words: comets: general - interplanetary medium - meteorites, meteors, mete- oroids - zodiacal dust 1 INTRODUCTION tention has been paid to the issue of episodes of bombard- ment, although several such have been noted and claims The discovery of an excess of iridium at the Cretaceous- have been made of correlation with mass extinction (Keller Tertiary boundary of 65 Myr BP, and its interpretation as et al 2012; McGhee 1996). -

Implications of a New Paleomagnetic Pole From
Latinmag Letters, Volume 3, Special Issue (2013), PB16, 1-6. Proceedings Montevideo, Uruguay THE PANGEA CONUNDRUM: IMPLICATIONS OF A NEW PALEOMAGNETIC POLE FROM THE PERMO-TRIASSIC ARAGUAINHA IMPACT STRUCTURE (CENTRAL BRAZIL) Elder Yokoyama1*, Daniele Brandt1, Eric Tohver2, Ricardo Trindade1 1 Instituto de Astronomia, Geofísica e Ciências Atmosféricas, Universidade de São Paulo, Rua do Matão, 1226, 05508-090, Brazil. 2 School of Earth and Environment, University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australia ABSTRACT The configuration of the Pangea supercontinent has been a topic of intense debate for almost half a century, a controversy that stems from discrepancies between the geology-based Pangea A and the paleomagnetically- based Pangea B. Recent paleomagnetic compilations aimed at resolving this controversy have identified the poor quality of paleomagnetic data from Gondwana for late Permian times as a major obstacle. Specifically, the vast majority of Gondwanan poles come from sedimentary rocks that are prone to biases from compaction or are poorly dated. Here we present a new paleomagnetic pole for cratonic South America based on impact- melts from the 254.7 ± 2.5 Ma Araguainha impact structure, an unusual target for paleomagnetic studies. The impact-generated melt sheet and veins were sampled at 22 sites (137 samples) and provide a reliable paleomagnetic record, similar to that of volcanic rocks. Alternating field and thermal demagnetization indicate stable, usually univectorial magnetizations carried by both magnetite and hematite. All sites but one show a single paleomagnetic direction of normal polarity with a mean direction of Dec = 356.4°; Inc = -38.7°; N = 21; k = 91.1; α95 = 3.3°, yielding a paleomagnetic pole (AIS) at Lat = -83.7; Lon = 340.1; K = 83.5; A95 =3.5°; SB =8.3°. -

The Geological Record of Meteorite Impacts
THE GEOLOGICAL RECORD OF METEORITE IMPACTS Gordon R. Osinski Canadian Space Agency, 6767 Route de l'Aeroport, St-Hubert, QC J3Y 8Y9 Canada, Email: [email protected] ABSTRACT 2. FORMATION OF METEORITE IMPACT STRUCTURES Meteorite impact structures are found on all planetary bodies in the Solar System with a solid The formation of hypervelocity impact craters has surface. On the Moon, Mercury, and much of Mars, been divided, somewhat arbitrarily, into three main impact craters are the dominant landform. On Earth, stages [3] (Fig. 2): (1) contact and compression, (2) 174 impact sites have been recognized, with several excavation, and (3) modification. A further stage of more new craters being discovered each year. The “hydrothermal and chemical alteration” is also terrestrial impact cratering record is critical for our considered as a separate, final stage in the cratering understanding of impacts as it currently provides the process (e.g., [4]), and is also described below. only ground-truth data on which to base interpretations of the cratering record of other planets and moons. In this contribution, I summarize the processes and products of impact cratering and provide and an up-to-date assessment of the geological record of meteorite impacts. 1. INTRODUCTION It is now widely recognized that impact cratering is a ubiquitous geological process that affects all planetary objects with a solid surface (e.g., [1]). One only has to look up on a clear night to see that impact structures are the dominant landform on the Moon. The same can be said of all the rocky and icy bodies in the solar system that have retained portions of their earliest crust. -

Geração, Mobilização E Cristalização De Um Fundido Rico Em Álcalis Na Estrutura De Impacto De Araguainha
Número 409/2009 UNIVERSIDADE ESTADUAL DE CAMPINAS INSTITUTO DE GEOCIÊNCIAS PÓS-GRADUAÇÃO EM GEOCIÊNCIAS UNICAMFI ÁREA DE GEOLOGIA E RECURSOS NATURAIS ROGÉRIO AMARO MACHADO GERAÇÃO, MOBILIZAÇÃO E CRISTALIZAÇÃO DE UM FUNDIDO RICO EM ÁLCALIS NA ESTRUTURA DE IMPACTO DE ARAGUAINHA Dissertação apresentada ao Instituto de Geociências como parte dos requisitos para a obtenção do título de Mestre em Geociências, Área de Geologia e Recursos Naturais Orientador: Pro f. Dr. Carlos Roberto de Souza Filho Co-orientador: Pro f Dr. Cristiano de Carvalho Lana CAMPINAS- SÃO PAULO Agosto - 2009 Catalogação na Publicação elaborada pela Biblioteca do Instituto de Geociências/UNICAMP ~· \~~ (\ Machado, Rogério Amaro ÁffiJ.0iü Geração, Mobilização e Cristalização de um Fundido Rico em Álcalis na Estrutura de Impacto de Araguainha I Rogério Amaro Machado.-- Campinas,SP.: [s.n.], 2009. Orientador: Carlos Roberto de Souza Filho. Co-Orientador: Cristiano de Carvalho Lana. Dissertação (mestrado) Universidade Estadual de Campinas, Instituto de Geociências. l. Estrutura de Impacto de Araguainha. 2. Fundidos de Impacto. 3. Geoquúnica. 4. Componente Meteorítica I. Souza Filho, Carlos Roberto de. Il. Lana, Cristiano de Carvalho. I. Universidade Estadual de Campinas, Instituto de Geociências. li. Universíty o f Stellenbosch, Centre for Clllstal Petrology, Department of(leo togy, Geography and Environmcnta! Stuclies. IIL Título. Título em inglês: Generation, Mobilization and Crystallization of an Alkali-Rich Melt on Araguainha Impact Structure. Palavras-chave em inglês (Keywords): Araguainha Impact Structure, Impact Melts, Geochemistry, Meteoritic Component. Área de concentração: Geologia e Recursos Naturais. Titulação: Mestre em Geociências. Banca examinadora: Carlos Roberto de Souza Filho; Lena Virgínia Soares Monteiro; Ruy Paulo Philipp. Data da defesa: 31108/2009. -

A Tale of Clusters: No Resolvable Periodicity in the Terrestrial Impact Cratering Record
Meier & Holm-Alwmark, 2017 – A tale of clusters (accepted for publication in MNRAS) A tale of clusters: No resolvable periodicity in the terrestrial impact cratering record Matthias M. M. Meier1* and Sanna Holm-Alwmark2 1ETH Zurich, Institute of Geochemistry & Petrology, Clausiusstrasse 25, 8092 Zurich, Switzerland 2Department of Geology, Lund University, S olvegatan 12, 22362 Lund, Sweden * E-Mail: [email protected] Accepted 2017 January 23. Received 2017 January 18; in original form 2016 July 09. This is a pre-copyedited, author-produced PDF of an article accepted for publication in Monthly Notices of the Royal Astronomical Society following peer review. The version of record is available at: dx.doi.org/10.1093/mnras/stx211 ABSTRACT Rampino & Caldeira (2015) carry out a circular spectral analysis (CSA) of the ter- restrial impact cratering record over the past 260 million years (Ma), and suggest a ~26 Ma periodicity of impact events. For some of the impacts in that analysis, new accurate and high-precision (“robust”; 2SE<2%) 40Ar-39Ar ages have recently been published, resulting in significant age shifts. In a CSA of the updated impact age list, the periodicity is strongly reduced. In a CSA of a list containing only im- pacts with robust ages, we find no significant periodicity for the last 500 Ma. We show that if we relax the assumption of a fully periodic impact record, assuming it to be a mix of a periodic and a random component instead, we should have found a periodic component if it contributes more than ~80% of the impacts in the last 260 Ma. -
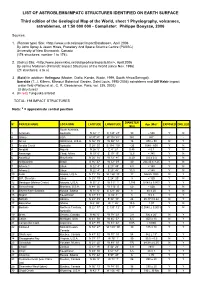
LIST of ASTROBLEMS/IMPATC STRUCTURES IDENTIFIED on EARTH SURFACE Third Edition of the Geological Map of the World, Sheet 1 Physi
LIST OF ASTROBLEMS/IMPATC STRUCTURES IDENTIFIED ON EARTH SURFACE Third edition of the Geological Map of the World, sheet 1 Physiography, volcanoes, astroblemes, at 1:50 000 000 - Compilator: Philippe Bouysse, 2006 Sources: 1. (Roman type) Site: <http://www.unb.ca/passc/ImpactDatabase>, April 2006 By John Spray & Jason Hines, Planetary And Space Science Centre (PASSC) University of New Brunswick, Canada (174 structures: number 1 to 174), 2. (Italics) Site: <http://www.somerikko.net/old/geo/imp/impacts.htm>, April 2006 By Jarmo Moilanen (Finland): Impact Structures of the World (since Nov. 1996) (21 structures: a to u) 3. (Bold) In addition: Velingara (Master, Diallo, Kande, Wade, 1999, South Africa/Senegal), Iturralde (T. J. Killeen, Missouri Botanical Garden, Saint Louis, 1995-2006) astroblems and Gilf Kebir impact crater field (Paillou et al., C. R. Géoscience, Paris, vol. 335, 2003). (3 structures) 4. (In red) Tunguska airblast TOTAL: 198 IMPACT STRUCTURES Nota: * = approximate central position DIAMETER N° CRATER NAME LOCATION LATITUDE LONGITUDE Age (Ma)* EXPOSED DRILLED (km) South Australia, 1 Acraman Australia S 32° 1' E 135° 27' 90 ~ 590 Y N a Alamo USA N 37°30' W 116°30' 190 367 - - 2 Ames Oklahoma, U.S.A. N 36° 15' W 98° 12' 16 470 ± 30 N Y 3 Amelia Creek Australia S 20° 55' E 134 ° 50' ~20 1640 - 600 Y N 4 Amguid Algeria N 26° 5' E 4° 23' 0.45 < 0.1 Y N 5 Aorounga Chad, Africa N 19° 6' E 19° 15' 12.6 < 345 Y N 6 Aouelloul Mauritania N 20° 15' W 12° 41' 0.39 3.0 ± 0.3 Y N 7 Araguainha Brazil S 16° 47' W 52° 59' 40 244.40 ± 3.25 Y N 8 Arkenu 1 Libya N 22° 4' E 23° 45' 6.8 < 140 Y N 9 Arkenu 2 Libya N 22° 4' E 23° 45' 10,3 < 140 Y N 10 Avak Alaska, U.S.A. -

Remote Sensing of Impact Craters 195
THIRTEEN Remote s ensing of i mpact c raters Shawn P. Wright * , Livio L. Tornabene † and Michael S. Ramsey ‡ * Institute of Meteoritics, MSC03 2050, University of New Mexico, Albuquerque, NM 87131, USA † Centre for Planetary Science and Exploration, Department of Earth Sciences Western University, 1151 Richmond Street, London, ON, N6A 5B7, Canada ‡ Department of Geology and Planetary Science, University of Pittsburgh, 4107 O ’ Hara Street, Pittsburgh, PA 15260 - 3332, USA 13.1 Introduction themes, followed by two case studies that utilize one or more of these aspects to solve a geological problem about an impact site. As discussed throughout this book, impact cratering is a funda- mental geological process that has shaped and modifi ed the surfaces of planets throughout geological time (see summary in 13.2 Background Chapter 1 ). However, the low frequency of meteorite impacts during the recent past and the ongoing active geological processes An early and ongoing goal involving remote sensing of terrestrial on Earth offer rare opportunities to examine pristine impact sites, impact craters has been to simply discover new impact craters which are common on the surfaces of other terrestrial planets. solely from remote - sensing data (Garvin et al ., 1992 ; Koeberl, With the exception of a few robotic and manned space explora- 2004 ; Folco et al ., 2010, 2011 ) or fi nd spectral differences between tion missions, the analysis of orbital data has been the primary shocked rocks and their protoliths (D ’ Aria and Garvin, 1988 ; method for studying impact craters in a variety of settings and Garvin et al ., 1992 ; Johnson et al ., 2002, 2007 ; Wright et al ., 2011 ). -

Structural Evolution of the 40 Km Wide Araguainha Impact Structure, Central Brazil
Meteoritics & Planetary Science 43, Nr 4, 701–716 (2008) Abstract available online at http://meteoritics.org Structural evolution of the 40 km wide Araguainha impact structure, central Brazil C. LANA1*, C. R. SOUZA FILHO2, Y. R. MARANGONI3, E. YOKOYAMA3, R. I. F. TRINDADE3, E. TOHVER4, and W. U. REIMOLD5 1Department of Geology, University of Stellenbosch, Private Bag X1, Matieland 7620, South Africa 2Departamento de Geologia e Recursos Naturais, Instituto de Geociencias, UNICAMP, 13083-970 Brazil 3Instituto de Astronomia, Geofísica e Ciencias Atmosfericas, USP Rua do Matao,1226, Cidade Universitaria, São Paulo, SP 05508-090, Brazil 4Tectonics Special Research Center, University of Western Australia, 35 Stirling Highway, Crawley, WA 6009, Australia 5Museum for Natural History (Mineralogy), Humboldt University, Invalidenstrasse 43, 10115 Berlin, Germany *Corresponding author. E-mail: [email protected] (Received 29 March 2007; revision accepted 05 September 2007) Abstract–The 40 km wide Araguainha structure in central Brazil is a shallowly eroded impact crater that presents unique insights into the final stages of complex crater formation. The dominant structural features preserved at Araguainha relate directly to the centripetal movement of the target rocks during the collapse of the transient cavity. Slumping of the transient cavity walls resulted in inward-verging inclined folds and a km-scale anticline in the outer ring of the structure. The folding stage was followed by radial and concentric faulting, with downward displacement of kilometer-scale blocks around the crater rim. The central uplift records evidence for km-scale upward movement of crystalline basement rocks from the transient cavity floor, and lateral moment of sedimentary target rocks detached from the cavity walls.