Antifungal and Phytotoxic Activities of Essential Oils: in Vitro Assays and Their Potential Use in Crop Protection
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Apiaceae) - Beds, Old Cambs, Hunts, Northants and Peterborough
CHECKLIST OF UMBELLIFERS (APIACEAE) - BEDS, OLD CAMBS, HUNTS, NORTHANTS AND PETERBOROUGH Scientific name Common Name Beds old Cambs Hunts Northants and P'boro Aegopodium podagraria Ground-elder common common common common Aethusa cynapium Fool's Parsley common common common common Ammi majus Bullwort very rare rare very rare very rare Ammi visnaga Toothpick-plant very rare very rare Anethum graveolens Dill very rare rare very rare Angelica archangelica Garden Angelica very rare very rare Angelica sylvestris Wild Angelica common frequent frequent common Anthriscus caucalis Bur Chervil occasional frequent occasional occasional Anthriscus cerefolium Garden Chervil extinct extinct extinct very rare Anthriscus sylvestris Cow Parsley common common common common Apium graveolens Wild Celery rare occasional very rare native ssp. Apium inundatum Lesser Marshwort very rare or extinct very rare extinct very rare Apium nodiflorum Fool's Water-cress common common common common Astrantia major Astrantia extinct very rare Berula erecta Lesser Water-parsnip occasional frequent occasional occasional x Beruladium procurrens Fool's Water-cress x Lesser very rare Water-parsnip Bunium bulbocastanum Great Pignut occasional very rare Bupleurum rotundifolium Thorow-wax extinct extinct extinct extinct Bupleurum subovatum False Thorow-wax very rare very rare very rare Bupleurum tenuissimum Slender Hare's-ear very rare extinct very rare or extinct Carum carvi Caraway very rare very rare very rare extinct Chaerophyllum temulum Rough Chervil common common common common Cicuta virosa Cowbane extinct extinct Conium maculatum Hemlock common common common common Conopodium majus Pignut frequent occasional occasional frequent Coriandrum sativum Coriander rare occasional very rare very rare Daucus carota Wild Carrot common common common common Eryngium campestre Field Eryngo very rare, prob. -

Chervil (Pronounced SHER-Vil) Is a Delicate Culinary Herb Used Frequently in French Cuisine
PO Box 218, Half Moon Bay, CA 94019 Phone: 650 726 4980 - Fax: 650 726 2991 Email: [email protected] To Grow a great Garden, Start With Amazing Seeds Chervil (pronounced SHER-vil) is a delicate culinary herb used frequently in French cuisine. It is a member of the parsley family with a mild flavor. This spring herb is often used in egg dishes. It is sometimes called French parsley. Chervil takes like a delicate cross between tarragon and parsley. Chervil has a mild flavor with hints of licorice or anise, but without those flavors coming through strongly. Chervil leaves are delicate and curly, somewhat resembling carrot greens. They are a little paler and have frillier leaves than flat-leaf parsley. Some bunches have tightly-closed leaves. You may find bunches that have blossoms, but you should avoid those because the herb will have turned bitter. Uses for Chervil…..Because of its delicate flavor, chervil is usually included in recipes, such as salads and soups It is added at the last minute as the flavor won't hold up in prolonged cooking; a good addition to omelets and is commonly used in making a classic Béarnaise sauce. It is included in the fines herbs blend, which is used in French cooking on poultry, egg dishes, and in salads. If you have chervil available, you might want to use it in herb infused oil, herb butter, or herb pesto to use with fish, poultry, eggs, soups, or salads. For example, use it in a recipe for roasted cod with wine and herb butter. -
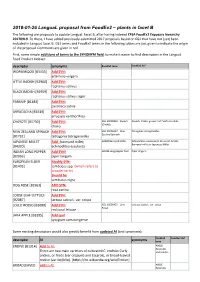
2018-01-26 Langual Proposal from Foodex2 – Plants in Facet B
2018-01-26 LanguaL proposal from FoodEx2 – plants in facet B The following are proposals to update LanguaL Facet B, after having indexed EFSA FoodEx2 Exposure hierarchy 20170919. To these, I have added previously-submitted 2017 proposals based on GS1 that have not (yet) been included in LanguaL facet B. GS1 terms and FoodEx2 terms in the following tables are just given to indicate the origin of the proposal. Comments are given in red. First, some simple additions of terms to the SYNONYM field, to make it easier to find descriptors in the LanguaL Food Product Indexer: descriptor synonyms FoodEx2 term FoodEx2 def WORMWOOD [B3433] Add SYN: artemisia vulgaris LITTLE RADISH [B2960] Add SYN: raphanus sativus BLACK RADISH [B2959] Add SYN: raphanus sativus niger PARSNIP [B1483] Add SYN: pastinaca sativa ARRACACHA [B3439] Add SYN: arracacia xanthorrhiza CHAYOTE [B1730] Add SYN: GS1 10006356 - Squash Squash, Choko, grown from Sechium edule (Choko) choko NEW ZEALAND SPINACH Add SYN: GS1 10006427 - New- Tetragonia tetragonoides Zealand Spinach [B1732] tetragonia tetragonoides JAPANESE MILLET Add : barnyard millet; A000Z Barnyard millet Echinochloa esculenta (A. Braun) H. Scholz, Barnyard millet or Japanese Millet. [B4320] echinochloa esculenta INDIAN LONG PEPPER Add SYN! A019B Long pepper fruit Piper longum [B2956] piper longum EUROPEAN ELDER Modify SYN: [B1403] sambucus spp. (which refers to broader term) Should be sambucus nigra DOG ROSE [B2961] ADD SYN: rosa canina LOOSE LEAF LETTUCE Add SYN: [B2087] lactusa sativa L. var. crispa LOLLO ROSSO [B2088] Add SYN: GS1 10006425 - Lollo Lactuca sativa L. var. crispa Rosso red coral lettuce JAVA APPLE [B3395] Add syn! syzygium samarangense Some existing descriptors would also greatly benefit from updated AI (and synonyms): FoodEx2 FoodEx2 def descriptor AI synonyms term ENDIVE [B1314] Add to AI: A00LD Escaroles There are two main varieties of cultivated C. -

Periodic Table of Herbs 'N Spices
Periodic Table of Herbs 'N Spices 11HH 1 H 2 HeHe Element Proton Element Symbol Number Chaste Tree Chile (Vitex agnus-castus) (Capsicum frutescens et al.) Hemptree, Agnus Cayenne pepper, Chili castus, Abraham's balm 118Uuo Red pepper 33LiLi 44 Be 5 B B 66 C 7 N 7N 88O O 99 F 1010 Ne Ne Picture Bear’s Garlic Boldo leaves Ceylon Cinnamon Oregano Lime (Allium ursinum) (Peumus boldus) (Cinnamomum zeylanicum) Nutmeg Origanum vulgare Fenugreek Lemon (Citrus aurantifolia) Ramson, Wild garlic Boldina, Baldina Sri Lanka cinnamon (Myristica fragrans) Oregan, Wild marjoram (Trigonella foenum-graecum) (Citrus limon) 11 Na Na 1212 Mg Mg 1313 Al Al 1414 Si Si 1515 P P 16 S S 1717 Cl Cl 1818 Ar Ar Common Name Scientific Name Nasturtium Alternate name(s) Allspice Sichuan Pepper et al. Grains of Paradise (Tropaeolum majus) (Pimenta dioica) (Zanthoxylum spp.) Perilla (Aframomum melegueta) Common nasturtium, Jamaica pepper, Myrtle Anise pepper, Chinese (Perilla frutescens) Guinea grains, Garden nasturtium, Mugwort pepper, Pimento, pepper, Japanese Beefsteak plant, Chinese Savory Cloves Melegueta pepper, Indian cress, Nasturtium (Artemisia vulgaris) Newspice pepper, et al. Basil, Wild sesame (Satureja hortensis) (Syzygium aromaticum) Alligator pepper 1919 K K 20 Ca Ca 2121 Sc Sc 2222 Ti Ti 23 V V 24 Cr Cr 2525 Mn Mn 2626 Fe Fe 2727 Co Co 2828 Ni Ni 29 Cu Cu 3030 Zn Zn 31 Ga Ga 3232 Ge Ge 3333As As 34 Se Se 3535 Br Br 36 Kr Kr Cassia Paprika Caraway (Cinnamomum cassia) Asafetida Coriander Nigella Cumin Gale Borage Kaffir Lime (Capsicum annuum) (Carum carvi) -

Herbs, Spices and Essential Oils
Printed in Austria V.05-91153—March 2006—300 Herbs, spices and essential oils Post-harvest operations in developing countries UNITED NATIONS INDUSTRIAL DEVELOPMENT ORGANIZATION Vienna International Centre, P.O. Box 300, 1400 Vienna, Austria Telephone: (+43-1) 26026-0, Fax: (+43-1) 26926-69 UNITED NATIONS FOOD AND AGRICULTURE E-mail: [email protected], Internet: http://www.unido.org INDUSTRIAL DEVELOPMENT ORGANIZATION OF THE ORGANIZATION UNITED NATIONS © UNIDO and FAO 2005 — First published 2005 All rights reserved. Reproduction and dissemination of material in this information product for educational or other non-commercial purposes are authorized without any prior written permission from the copyright holders provided the source is fully acknowledged. Reproduction of material in this information product for resale or other commercial purposes is prohibited without written permission of the copyright holders. Applications for such permission should be addressed to: - the Director, Agro-Industries and Sectoral Support Branch, UNIDO, Vienna International Centre, P.O. Box 300, 1400 Vienna, Austria or by e-mail to [email protected] - the Chief, Publishing Management Service, Information Division, FAO, Viale delle Terme di Caracalla, 00100 Rome, Italy or by e-mail to [email protected] The designations employed and the presentation of material in this information product do not imply the expression of any opinion whatsoever on the part of the United Nations Industrial Development Organization or of the Food and Agriculture Organization of the United Nations concerning the legal or development status of any country, territory, city or area or of its authorities, or concerning the delimitation of its frontiers or boundaries. -

Spice Basics
SSpicepice BasicsBasics AAllspicellspice Allspice has a pleasantly warm, fragrant aroma. The name refl ects the pungent taste, which resembles a peppery compound of cloves, cinnamon and nutmeg or mace. Good with eggplant, most fruit, pumpkins and other squashes, sweet potatoes and other root vegetables. Combines well with chili, cloves, coriander, garlic, ginger, mace, mustard, pepper, rosemary and thyme. AAnisenise The aroma and taste of the seeds are sweet, licorice like, warm, and fruity, but Indian anise can have the same fragrant, sweet, licorice notes, with mild peppery undertones. The seeds are more subtly fl avored than fennel or star anise. Good with apples, chestnuts, fi gs, fi sh and seafood, nuts, pumpkin and root vegetables. Combines well with allspice, cardamom, cinnamon, cloves, cumin, fennel, garlic, nutmeg, pepper and star anise. BBasilasil Sweet basil has a complex sweet, spicy aroma with notes of clove and anise. The fl avor is warming, peppery and clove-like with underlying mint and anise tones. Essential to pesto and pistou. Good with corn, cream cheese, eggplant, eggs, lemon, mozzarella, cheese, olives, pasta, peas, pizza, potatoes, rice, tomatoes, white beans and zucchini. Combines well with capers, chives, cilantro, garlic, marjoram, oregano, mint, parsley, rosemary and thyme. BBayay LLeafeaf Bay has a sweet, balsamic aroma with notes of nutmeg and camphor and a cooling astringency. Fresh leaves are slightly bitter, but the bitterness fades if you keep them for a day or two. Fully dried leaves have a potent fl avor and are best when dried only recently. Good with beef, chestnuts, chicken, citrus fruits, fi sh, game, lamb, lentils, rice, tomatoes, white beans. -

Angelica Cornflowers Lemon Balm Verbena Salad Burnet Savory Tarragon Chervil Nasturtium Pansy Oregano Sage Fennel Dill Borage Ma
Summertime herbs and edible flowers Angelica Borage Chervil Cornflowers The shiny dark green leaves of ornamental Borage has a refreshing, cooling, slightly bitter, Chervil is a mild herb and a generous quantity of The petals of cornflowers can be plucked and angelica pachycarpa make a wonderful garnish. cucumber flavour. The light purple flowers are chopped leaves is best added fresh just before added to salads. Blue cornflowers are most Another type is angelica archangelica, the culinary most commonly used as a garnish on salads or serving as the light flavour is quickly lost. It can common although pink and white cornflowers and medicinal plant. It has matt green leaves desserts.The small young leaves can be used be used in all sorts of salads, or sprinkled over are also edible. which wilt quickly. The stems are crystallised for in salads or lightly cooked with other green lightlycooked vegetables or soups. Chervil is a garnish on cakes and desserts, and the root is vegetables. The flowers can be set in ice cubes much like parsley but is more subtle; it goes well used for medicinal purposes. or crystallised. with egg dishes, meats, poultry and fish. Marigold Marigold flowers, very similar to calendula, make an attractive edible garnish or they can be added Dill sparingly to salads. The petals can be used in place of saffron and will give colour in many Dill leaves and seeds have a mild aniseed flavour Fennel Lemon balm dishes, especially rice and egg dishes. which is similar though slightly sweeter and more Fennel grows wild by the roadside and may even Lemon balm leaves give a delicate lemon sweet aromatic than fennel. -

Taxonomy, Origin and Importance of the Apiaceae Family
1 TAXONOMY, ORIGIN AND IMPORTANCE OF THE APIACEAE FAMILY JEAN-PIERRE REDURON* Mulhouse, France The Apiaceae (or Umbelliferae) is a plant family comprising at the present time 466 genera and about 3800 species (Plunkett et al., 2018). It is distributed nearly worldwide, but is most diverse in temperate climatic areas, such as Eurasia and North America. It is quite rare in tropical humid regions where it is limited to high mountains. Mediterranean and arid climatic conditions favour high species diversification. The Apiaceae are present in nearly all types of habi- tats, from sea-level to alpine zones: aquatic biotopes, grasslands, grazed pas- tures, forests including their clearings and margins, cliffs, screes, rocky hills, open sandy and gravelly soils, steppes, cultivated fields, fallows, road sides and waste grounds. The largest number of genera, 289, and the largest generic endemism, 177, is found in Asia. There are 126 genera in Europe, but only 17 are en- demic. Africa has about the same total with 121 genera, where North Africa encompasses the largest occurrence of 82 genera, 13 of which are endemic. North and Central America have a fairly high level of diversity with 80 genera and 44 endemics, where South America accommodates less generic diversity with 35 genera, 15 of which are endemic. Oceania is home to 27 genera and 18 endemics (Plunkett et al., 2018). The Apiaceae family appears to have originated in Australasia (region including Australia, Tasmania, New Zealand, New Guinea, New Caledonia and several island groups), with this origin dated to the Late Cretaceous/ early Eocene, c.87 Ma (Nicolas and Plunkett, 2014). -

Wild Chervil Factsheet
K. Weller FACTSHEET APRIL 2019 Wild Chervil Anthriscus sylvestris About Wild Chervil Native to Europe, Wild Chervil is thought to have been introduced through wildflower seed mixes. It will take over many habitats, but particularly thrives in wet, disturbed soil. It is very difficult to remove because of its long taproot and tendency to grow near bodies of water, which make many herbicides unusable. Wild Chervil competes with native plants species and hay crops, harming both the economy and environment. Legal Status Noxious Weed (Regional), BC Weed Control Act S. Dewey, Bugwood.org Fruits: Each flower produces two joined seeds about 6-7mm in length. They are smooth, shiny and dark brown. Similar Species: There are a few species that can easily be confused with Wild chervil. Some include Bur Chervil (Anthriscus caucalis), Rough Chervil (Chaerophyllum temulum) Wild chervil Daucus carota Anthriscus sylvestris and Queen Anne’s lace ( ). Bur chervil can be Code = WI n = 1246 IAPP July 2017 easily distinguished from Wild chervil by its rounder and lighter Sites per 10km block squares 1-10 green leaves. Rough Chervil can be distinguished by the purple 11-20 spots on its stem. Queen Anne’s lace can be distinguished by its 21-50 unique reddish-purple flower found in the center of its umbel of 51-100 white flowers. Over 100 0 200km 400km Ecological Characteristics Distribution Habitat: Prefers wet to moist disturbed sites such as pastures, fields, roadsides, and fence lines. Will grow in a variety of soils but Wild Chervil is mostly distributed throughout the southern prefers low to mid elevation. -

Wild Chervil (Anthriscus Sylvestris) Is an Invasive Biennial Or Short-Lived Perennial Plant of the Carrot Family
Wild Chervil (Anthriscus sylvestris) is an invasive biennial or short-lived perennial plant of the carrot family. The plant was introduced from Eurasia, and is becoming widely naturalized in eastern Canada where it is a weed of perennial forage crops and pastures. It is a noxious weed in Nova Scotia under the Agricultural Weed Control Act Wildand Weed Chervil Control Regulations. How to Identify Wild Chervil Habitat and Characteristics A rosette of leaves in its first year, with a flowering of Wild Chervil stem up to 1.5 meters in later years. Looks similar Grows in a variety of soil types, found in ditches, to Queen Anne’s Lace, Caraway, and Sweet Cicely. dykes, roadsides, meadows, pasture, perennial Leaves Pinnately compound, triangular in shape, forage fields, stream banks and hedge rows. fern-like in appearance. Leaves alternate along Spreads by seed, forming dense patches the stem with the base clasping along the through root budding. stem as well. Slightly hairy underneath. Aggressively competes for light, space, Stem Grooved, branched, hollow, nutrients and water, often crowding covered in small hairs. out desirable vegetation. Flowers Small white flowers Sometimes grazed, it has little in large compound umbels, nutritional value as a forage for blooming from, late May livestock. If harvested in forage, through July. it can reduce crop quality due to mold. Seeds occur in pairs, greenish brown Wild Chervil also serves as a host for to dark brown, smooth and shiny, the parsnip yellow fleck virus, which elongated oval in shape. 5–10mm long, can infect carrots, celery and parsnips. cylindrical and tapering to a beak. -

Dictionary of Cultivated Plants and Their Regions of Diversity Second Edition Revised Of: A.C
Dictionary of cultivated plants and their regions of diversity Second edition revised of: A.C. Zeven and P.M. Zhukovsky, 1975, Dictionary of cultivated plants and their centres of diversity 'N -'\:K 1~ Li Dictionary of cultivated plants and their regions of diversity Excluding most ornamentals, forest trees and lower plants A.C. Zeven andJ.M.J, de Wet K pudoc Centre for Agricultural Publishing and Documentation Wageningen - 1982 ~T—^/-/- /+<>?- •/ CIP-GEGEVENS Zeven, A.C. Dictionary ofcultivate d plants andthei rregion so f diversity: excluding mostornamentals ,fores t treesan d lowerplant s/ A.C .Zeve n andJ.M.J ,d eWet .- Wageninge n : Pudoc. -11 1 Herz,uitg . van:Dictionar y of cultivatedplant s andthei r centreso fdiversit y /A.C .Zeve n andP.M . Zhukovsky, 1975.- Me t index,lit .opg . ISBN 90-220-0785-5 SISO63 2UD C63 3 Trefw.:plantenteelt . ISBN 90-220-0785-5 ©Centre forAgricultura l Publishing and Documentation, Wageningen,1982 . Nopar t of thisboo k mayb e reproduced andpublishe d in any form,b y print, photoprint,microfil m or any othermean swithou t written permission from thepublisher . Contents Preface 7 History of thewor k 8 Origins of agriculture anddomesticatio n ofplant s Cradles of agriculture and regions of diversity 21 1 Chinese-Japanese Region 32 2 Indochinese-IndonesianRegio n 48 3 Australian Region 65 4 Hindustani Region 70 5 Central AsianRegio n 81 6 NearEaster n Region 87 7 Mediterranean Region 103 8 African Region 121 9 European-Siberian Region 148 10 South American Region 164 11 CentralAmerica n andMexica n Region 185 12 NorthAmerica n Region 199 Specieswithou t an identified region 207 References 209 Indexo fbotanica l names 228 Preface The aimo f thiswor k ist ogiv e thereade r quick reference toth e regionso f diversity ofcultivate d plants.Fo r important crops,region so fdiversit y of related wild species areals opresented .Wil d species areofte nusefu l sources of genes to improve thevalu eo fcrops . -

Epicentre Spice Guide
SPICE The Epicentre Guidebook Edited by Barry Mortin © 2013, The Epicentre. All rights reserved Please Note: This ebook is by no means a complete encyclopedia of spices. Please refer to our website, theepicentre.com for more comprehensive information. Please register for updates, special offers and to win spices! Register at theepicentre.com SPICE The Epicentre Guidebook CONTENTS Ajwain .....................................................................1 Herbes de Provence.........................................43 Aleppo Pepper.....................................................2 Juniper..................................................................44 Allspice ...................................................................4 Kaffir Lime leaves..............................................46 Amchur...................................................................7 Lavender..............................................................48 Ancho Chile...........................................................8 Lemongrass .......................................................50 Anise .......................................................................9 Mace......................................................................51 Annatto................................................................11 Mint .......................................................................53 Basil .......................................................................13 Nigella ..................................................................55