The Potential of Planets Orbiting Red Dwarf Stars to Support Oxygenic Photosynthesis and Complex Life
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-
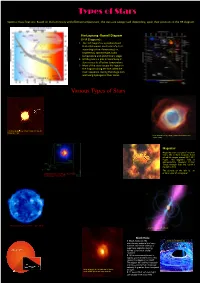
Brown Dwarf: White Dwarf: Hertzsprung -Russell Diagram (H-R
Types of Stars Spectral Classifications: Based on the luminosity and effective temperature , the stars are categorized depending upon their positions in the HR diagram. Hertzsprung -Russell Diagram (H-R Diagram) : 1. The H-R Diagram is a graphical tool that astronomers use to classify stars according to their luminosity (i.e. brightness), spectral type, color, temperature and evolutionary stage. 2. HR diagram is a plot of luminosity of stars versus its effective temperature. 3. Most of the stars occupy the region in the diagram along the line called the main sequence. During that stage stars are fusing hydrogen in their cores. Various Types of Stars Brown Dwarf: White Dwarf: Brown dwarfs are sub-stellar objects After a star like the sun exhausts its nuclear that are not massive enough to sustain fuel, it loses its outer layer as a "planetary nuclear fusion processes. nebula" and leaves behind the remnant "white Since, comparatively they are very cold dwarf" core. objects, it is difficult to detect them. Stars with initial masses Now there are ongoing efforts to study M < 8Msun will end as white dwarfs. them in infrared wavelengths. A typical white dwarf is about the size of the This picture shows a brown dwarf around Earth. a star HD3651 located 36Ly away in It is very dense and hot. A spoonful of white constellation of Pisces. dwarf material on Earth would weigh as much as First directly detected Brown Dwarf HD 3651B. few tons. Image by: ESO The image is of Helix nebula towards constellation of Aquarius hosts a White Dwarf Helix Nebula 6500Ly away. -

A FEROS Survey of Hot Subdwarf Stars
Open Astron. 2018; 27: 7–13 Research Article Stéphane Vennes*, Péter Németh, and Adela Kawka A FEROS Survey of Hot Subdwarf Stars https://doi.org/10.1515/astro-2018-0005 Received Oct 02, 2017; accepted Nov 07, 2017 Abstract: We have completed a survey of twenty-two ultraviolet-selected hot subdwarfs using the Fiber-fed Extended Range Optical Spectrograph (FEROS) and the 2.2-m telescope at La Silla. The sample includes apparently single objects as well as hot subdwarfs paired with a bright, unresolved companion. The sample was extracted from our GALEX cat- alogue of hot subdwarf stars. We identified three new short-period systems (P = 3.5 hours to 5 days) and determined the orbital parameters of a long-period (P = 62d.66) sdO plus G III system. This particular system should evolve into a close double degenerate system following a second common envelope phase. We also conducted a chemical abundance study of the subdwarfs: Some objects show nitrogen and argon abundance excess with respect to oxygen. We present key results of this programme. Keywords: binaries: close, binaries: spectroscopic, subdwarfs, white dwarfs, ultraviolet: stars 1 Introduction ing a mass transfer event, i.e., comparable to the observed fraction (≈70%) of short period binaries (P < 10 d, Maxted et al. 2001; Morales-Rueda et al. 2003), while the remain- The properties of extreme horizontal branch (EHB) stars, ing single objects are formed by the merger of two helium i.e., the hot, hydrogen-rich (sdB) and helium-rich subd- white dwarfs which would also result in a thin hydrogen warf (sdO) stars, located at the faint blue end of the hori- layer. -

Massive Fast Rotating Highly Magnetized White Dwarfs: Theory and Astrophysical Applications
Massive Fast Rotating Highly Magnetized White Dwarfs: Theory and Astrophysical Applications Thesis Advisors Ph.D. Student Prof. Remo Ruffini Diego Leonardo Caceres Uribe* Dr. Jorge A. Rueda *D.L.C.U. acknowledges the financial support by the International Relativistic Astrophysics (IRAP) Ph.D. program. Academic Year 2016–2017 2 Contents General introduction 4 1 Anomalous X-ray pulsars and Soft Gamma-ray repeaters: A new class of pulsars 9 2 Structure and Stability of non-magnetic White Dwarfs 21 2.1 Introduction . 21 2.2 Structure and Stability of non-rotating non-magnetic white dwarfs 23 2.2.1 Inverse b-decay . 29 2.2.2 General Relativity instability . 31 2.2.3 Mass-radius and mass-central density relations . 32 2.3 Uniformly rotating white dwarfs . 37 2.3.1 The Mass-shedding limit . 38 2.3.2 Secular Instability in rotating and general relativistic con- figurations . 38 2.3.3 Pycnonuclear Reactions . 39 2.3.4 Mass-radius and mass-central density relations . 41 3 Magnetic white dwarfs: Stability and observations 47 3.1 Introduction . 47 3.2 Observations of magnetic white dwarfs . 49 3.2.1 Introduction . 49 3.2.2 Historical background . 51 3.2.3 Mass distribution of magnetic white dwarfs . 53 3.2.4 Spin periods of isolated magnetic white dwarfs . 53 3.2.5 The origin of the magnetic field . 55 3.2.6 Applications . 56 3.2.7 Conclusions . 57 3.3 Stability of Magnetic White Dwarfs . 59 3.3.1 Introduction . 59 3.3.2 Ultra-magnetic white dwarfs . 60 3.3.3 Equation of state and virial theorem violation . -

Asymptotic Giant Branch Helium Flash Horizontal Branch
The Death of Stars - I. Learning Objectives ! Be able to sketch the H-R diagram and include stars by size, spectral type, lifetime, color, mass, magnitude, temperature and luminosity, relative to our Sun ! Compare Red Dwarfs to our Sun in terms of their different masses and interiors ! Know the physics of the evolution of low mass stars from Main Sequence to white dwarf (fusion, gravity, collapse, expansion, temperature, composition, equilibrium) ! How do evolving stars move around the H-R diagram (where do they go when expanding, helium burning etc?) ! How does electron degeneracy pressure stabilize a dead star? What is a white dwarf? What is a planetary nebula? Important bad drawing So much information in the HR diagram... The Evolution of Stars ! A star’s evolution depends on its mass ! We will look at the evolution of three general types of stars ! Red dwarf stars (less than 0.4 M⊙ ) ! Low mass stars (0.4 to 8 M⊙ ) ! High mass stars (more than 8 M⊙ ) ! We can track the evolution of a star on the H-R diagram ! From Main Sequence to giant/supergiant and to its final demise Red Dwarf Stars ! 0.08 M⊙ < Mass < 0.4 M⊙ ! Fully convective interior (no “radiative zone”) ! Burn hydrogen slowly as convection constantly circulates hydrogen to the core ! Live 100s of billions to trillions of years ! The Universe is only 13.8 billion years old, so none of these stars have died yet Stellar Demise! Low mass star Helium-burning White dwarf and red giant planetary nebula High mass Helium-burning Other supergiant Core-collapse star red supergiant phases -

Index to JRASC Volumes 61-90 (PDF)
THE ROYAL ASTRONOMICAL SOCIETY OF CANADA GENERAL INDEX to the JOURNAL 1967–1996 Volumes 61 to 90 inclusive (including the NATIONAL NEWSLETTER, NATIONAL NEWSLETTER/BULLETIN, and BULLETIN) Compiled by Beverly Miskolczi and David Turner* * Editor of the Journal 1994–2000 Layout and Production by David Lane Published by and Copyright 2002 by The Royal Astronomical Society of Canada 136 Dupont Street Toronto, Ontario, M5R 1V2 Canada www.rasc.ca — [email protected] Table of Contents Preface ....................................................................................2 Volume Number Reference ...................................................3 Subject Index Reference ........................................................4 Subject Index ..........................................................................7 Author Index ..................................................................... 121 Abstracts of Papers Presented at Annual Meetings of the National Committee for Canada of the I.A.U. (1967–1970) and Canadian Astronomical Society (1971–1996) .......................................................................168 Abstracts of Papers Presented at the Annual General Assembly of the Royal Astronomical Society of Canada (1969–1996) ...........................................................207 JRASC Index (1967-1996) Page 1 PREFACE The last cumulative Index to the Journal, published in 1971, was compiled by Ruth J. Northcott and assembled for publication by Helen Sawyer Hogg. It included all articles published in the Journal during the interval 1932–1966, Volumes 26–60. In the intervening years the Journal has undergone a variety of changes. In 1970 the National Newsletter was published along with the Journal, being bound with the regular pages of the Journal. In 1978 the National Newsletter was physically separated but still included with the Journal, and in 1989 it became simply the Newsletter/Bulletin and in 1991 the Bulletin. That continued until the eventual merger of the two publications into the new Journal in 1997. -
Lecture 15: Stars
Matthew Schwartz Statistical Mechanics, Spring 2019 Lecture 15: Stars 1 Introduction There are at least 100 billion stars in the Milky Way. Not everything in the night sky is a star there are also planets and moons as well as nebula (cloudy objects including distant galaxies, clusters of stars, and regions of gas) but it's mostly stars. These stars are almost all just points with no apparent angular size even when zoomed in with our best telescopes. An exception is Betelgeuse (Orion's shoulder). Betelgeuse is a red supergiant 1000 times wider than the sun. Even it only has an angular size of 50 milliarcseconds: the size of an ant on the Prudential Building as seen from Harvard square. So stars are basically points and everything we know about them experimentally comes from measuring light coming in from those points. Since stars are pointlike, there is not too much we can determine about them from direct measurement. Stars are hot and emit light consistent with a blackbody spectrum from which we can extract their surface temperature Ts. We can also measure how bright the star is, as viewed from earth . For many stars (but not all), we can also gure out how far away they are by a variety of means, such as parallax measurements.1 Correcting the brightness as viewed from earth by the distance gives the intrinsic luminosity, L, which is the same as the power emitted in photons by the star. We cannot easily measure the mass of a star in isolation. However, stars often come close enough to another star that they orbit each other. -

A Ds9 Activity
Analyzing X-Ray Pulses from Stellar Cores – Pencil & Paper Version Purpose: To determine if two end products of stellar evolution – GK Per and Cen X-3 – could be white dwarfs or neutron stars by calculating the periods of X-ray pulses emitted by the two stellar cores using data recorded by the Chandra X-ray Observatory. Background: Variable Stars are stars that vary in magnitude (brightness). The behavior of stars that vary in magnitude can be studied by measuring their changes in magnitude over time and plotting the changes on a SS Cygni Light Curve, AAVSO graph called a light curve. The example to the left is a light curve that shows the variability of SS Cygni over a 300 day period of observation. SS Cygni is a cataclysmic variable – stars that have occasional violent outbursts caused by thermonuclear processes either in their surface layers or deep within their interiors. SS Cygni is also an example of a subclass of cataclysmic variables called dwarf novae. This is a close binary system consisting of a red dwarf star, cooler and less massive than the Sun, a white dwarf, and an accretion disk surrounding the white dwarf. The brightening of this system occurs when instability within the disk forces some of the disk material to drain down onto the surface of the white dwarf triggering a Illustration by David T Hardy, Artist thermonuclear explosion. The SS Cygni light curve above shows 5 thermonuclear events within the 300 day observation period. The SS Cygni light curve above was observed in the optical part of the spectrum and displays the dynamics among the accretion disk – the material in the disk that was accreted from its companion star – and the surface of the white dwarf. -

Optical Studies of X-Ray Sources in the Old Open Cluster M 67
Optical studies of X-ray sources in the old open cluster M 67 Figures 1.1 and 8.1, the images in the chapter headings and the images at the end are created from the Digitized Sky Survey c 2001 Maureen van den Berg Alle rechten voorbehouden. Niets in deze uitgave mag worden verveelvoudigd, opgesla- gen in een geautomatiseerd gegevensbestand, of openbaar gemaakt, in enige vorm, zonder schriftelijke toestemming van de auteur ISBN 90-393-2679-7 Optical studies of X-ray sources in the old open cluster M 67 Optische studies van rontgenbronnen¨ in de oude open sterrenhoop M 67 (met een samenvatting in het Nederlands) Proefschrift ter verkrijging van de graad van doctor aan de Universiteit Utrecht op gezag van de Rector Magnificus prof. dr H.O. Voorma ingevolge het besluit van het College voor Promoties, in het openbaar te verdedigen op woensdag 21 maart 2001 des ochtends te 10.30 uur door Maureen Constance van den Berg geboren op 6 april 1972 te Maastricht PROMOTOR prof. dr Frank Verbunt Dit proefschrift werd mede mogelijk gemaakt met financiele¨ steun van de Nederlandse Organisatie voor Wetenschappelijk Onderzoek (NWO). Contents 1 Introduction 1 1.1 Blue stragglers, yellow stragglers and sub-subgiants 1 1.2 X-ray observations of old open clusters 3 1.3 Summary: peculiar X-ray sources in M 67, NGC 752 and NGC 6940 5 2 Optical spectroscopy of X-ray sources in the old open cluster M67 9 2.1 Introduction 10 2.2 Observations and data reduction 12 2.2.1 Low-resolution spectra 14 2.2.2 High-resolution spectra 14 2.3 Data analysis 15 2.3.1 Determination of Ca II H&K emission fluxes 15 2.3.2 Determination of projected rotational velocities 17 2.4 Results 19 2.4.1 Comparison with RS CVn binaries 19 2.4.2 Activity indicators 21 2.4.3 Individual systems 22 2.5 Discussion and conclusions 26 3 Photometric variability in the old open cluster M 67 I. -
Solar-Type Activity in Main-Sequence Stars.Pdf
ASTRONOMY AND ASTROPHYSICS LIBRARY Series Editors: G. B¨orner, Garching, Germany A. Burkert, M¨unchen, Germany W. B. Burton, Charlottesville, VA, USA and Leiden, The Netherlands M.A. Dopita, Canberra, Australia A. Eckart, K¨oln, Germany T. Encrenaz, Meudon, France M. Harwit, Washington, DC, USA R. Kippenhahn, G¨ottingen, Germany B. Leibundgut, Garching, Germany J. Lequeux, Paris, France A. Maeder, Sauverny, Switzerland V. Trimble, College Park, MD, and Irvine, CA, USA R. E. Gershberg Solar-Type Activity in Main-Sequence Stars Translated by S. Knyazeva With 69 Figures and 17 Tables 123 Professor Roald E. Gershberg Crimean Astrophysical Observatory Crimea Nauchny 98409, Ukraine Dr. Svetlana Knyazeva Department of International Programmes Siberian Branch of RAS 17, Prosp. Akademika Lavrentieva Novosibirsk 630090, Russia Cover picture: Flare on AD Leo of 18 May 1965. (Gershberg and Chugainov, 1966) Library of Congress Control Number: 2005926088 ISSN 0941-7834 ISBN-10 3-540-21244-2 Springer Berlin Heidelberg New York ISBN-13 978-3-540-21244-7 Springer Berlin Heidelberg New York This work is subject to copyright. All rights are reserved, whether the whole or part of the material is concerned, specifically the rights of translation, reprinting, reuse of illustrations, recitation, broadcasting, reproduction on mi- crofilm or in any other way, and storage in data banks. Duplication of this publication or parts thereof is permitted only under the provisions of the German Copyright Law of September 9, 1965, in its current version, and permission for use must always be obtained from Springer. Violations are liable to prosecution under the German Copyright Law. Springer is a part of Springer Science+Business Media springeronline.com © Springer Berlin Heidelberg 2005 Printed in The Netherlands The use of general descriptive names, registered names, trademarks, etc. -

Lecture 16: Evolution of Low-Mass Stars Readings: 21-1, 21-2, 22-1, 22-3 and 22-4
Lecture 16: Evolution of Low-Mass Stars Readings: 21-1, 21-2, 22-1, 22-3 and 22-4 For the protostar and pre-main-sequence phases, the process was the same for the high and low mass stars, and the main difference was the speed with which they went through the various stages. On the main sequence, high and low mass stars are again quite similar. They fuse H to He in their cores, have cooler envelopes, and are in hydrostatic and thermal equilibrium. There are some differences: energy transport, CNO vs. proton-proton and lifetime on the main-sequence, but both high and low mass stars pass through this phase. However, once stars leave the main-sequence, high and low mass stars have very different paths. And the key difference is whether they end their lives as white dwarfs or as neutron stars/black holes. We look first at the evolution of low-mass stars after the main-sequence Key Ideas Low Mass Star = M < 8 Msun Stages of Evolution of a Low Mass Star Main Sequence star Red Giant star Horizontal Branch star Asymptotic Giant Branch star Planetary Nebula phase White Dwarf Please note that after the Red Giant phase, the names of the phases are exceedingly unhelpful and the result of history. But you’ll need to learn them nonetheless. Main Sequence Phase Energy Source: H fusion in the core What happens to the He from H fusion? Too cool to ignite He fusion Slowly build up an inert He core M-S Lifetime ~10 Gyr for a 1 MSun ~10 Tyr for a 0.1 MSun (red dwarf or M dwarf) Hydrogen Exhaustion Inside: Loss of pressure=end of hydrostatic equilibrium He -

Searching the Skies
SEARCHING THE SKIES THE UNIVERSE IS FILLED WITH ONE BILLION TRILLION STARS – LOOK AT THE SKY ON A CLEAR NIGHT AND THE FEW THOUSAND THAT ARE VISIBLE ARE INCREDIBLE TO SEE. THE DR BRAD NEWTON FASCINATION OF GAZING INTO THE VASTNESS OF THE UNIVERSE HAS LED IN THE US, TO DELVE BARLOW, OF THE CULP PLANETARIUM AND HIGH POINT UNIVERSITY DEEPER INTO SPACE TO HUNT FOR A STRANGE TYPE OF STAR CALLED A HOT SUBDWARF TALK LIKE AN ASTROPHYSICIST subdwarf merges with its neighbouring white dwarf, it may explode to form a Type 1a supernova. Cosmologists use these supernovae ASTRONOMY – the study of the Universe RED GIANT – a star that has expanded to measure vast distances across the Universe to a tremendous diameter during the final and have shown that the expansion of the ASTROPHYSICS – a branch of astronomy stages of its lifetime Universe is accelerating. We frequently which studies the physical processes and observe Type 1a supernovae explosions in other properties of the Universe TYPE 1A SUPERNOVA – the explosion of galaxies, but astronomers have not caught a white dwarf upon exceeding its mass limit a binary system just before it collides and BINARY STAR SYSTEM – two stars explodes. Until now… which orbit each other VARIABLE STAR – a star whose brightness changes as we view it from Earth Brad helped discover a new binary system COSMOLOGY – a branch of astronomy containing two stars so close together that which studies the origin and evolution of the WHITE DWARF – the extremely dense, they orbit each other in under two hours. Universe Earth-sized remnant core of a dead star “Earth takes 365 days to orbit the sun,” explains Brad, “but these two stars orbit PYTHON – a scripting language used in in under two hours!” They are so close and computer coding orbiting so quickly that they are slowly spiralling in. -

Galaxies and Stars
Galaxies and Stars 1. To an observer on Earth, the Sun appears brighter than the star Rigel because the Sun is A) hotter than Rigel B) more luminous than Rigel C) closer than Rigel D) larger than Rigel 2. Light and other forms of electromagnetic radiation are given off by stars using energy released during A) nuclear fusion B) conduction C) convection D) radioactive decay 3. Base your answer to the following question on The reaction below represents an energy-producing process. The reaction represents how energy is produced A) in the Sun by fusion B) when water condenses in Earth’s atmosphere C) from the movement of crustal plates D) during nuclear decay 4. The reaction below represents an energy-producing process. The reaction represents how energy is produced A) in the Sun by fusion B) when water condenses in Earth’s atmosphere C) from the movement of crustal plates D) during nuclear decay Base your answers to questions 5 and 6 on the graph below, which shows changes in the Sun’s magnetic activity and changes in the number of sunspots over a period of approximately 100 years. Sunspots are dark, cooler areas within the Sun’s photosphere that can be seen from Earth. 5. Which graph best represents the relationship between the number of sunspots and the amount of magnetic activity in the Sun? A) B) C) D) 6. The graph indicates that years having the greatest number of sunspots occur A) randomly and unpredictably B) precisely at the beginning of each decade C) in a cyclic pattern, repeating approximately every 6 years D) in a cyclic pattern, repeating approximately every 11 years 7.