Recent Collections of Fishes at the Kermadec Islands and New Records for the Region
Total Page:16
File Type:pdf, Size:1020Kb
Load more
Recommended publications
-

Biodiversity of the Kermadec Islands and Offshore Waters of the Kermadec Ridge: Report of a Coastal, Marine Mammal and Deep-Sea Survey (TAN1612)
Biodiversity of the Kermadec Islands and offshore waters of the Kermadec Ridge: report of a coastal, marine mammal and deep-sea survey (TAN1612) New Zealand Aquatic Environment and Biodiversity Report No. 179 Clark, M.R.; Trnski, T.; Constantine, R.; Aguirre, J.D.; Barker, J.; Betty, E.; Bowden, D.A.; Connell, A.; Duffy, C.; George, S.; Hannam, S.; Liggins, L..; Middleton, C.; Mills, S.; Pallentin, A.; Riekkola, L.; Sampey, A.; Sewell, M.; Spong, K.; Stewart, A.; Stewart, R.; Struthers, C.; van Oosterom, L. ISSN 1179-6480 (online) ISSN 1176-9440 (print) ISBN 978-1-77665-481-9 (online) ISBN 978-1-77665-482-6 (print) January 2017 Requests for further copies should be directed to: Publications Logistics Officer Ministry for Primary Industries PO Box 2526 WELLINGTON 6140 Email: [email protected] Telephone: 0800 00 83 33 Facsimile: 04-894 0300 This publication is also available on the Ministry for Primary Industries websites at: http://www.mpi.govt.nz/news-resources/publications.aspx http://fs.fish.govt.nz go to Document library/Research reports © Crown Copyright - Ministry for Primary Industries TABLE OF CONTENTS EXECUTIVE SUMMARY 1 1. INTRODUCTION 3 1.1 Objectives: 3 1.2 Objective 1: Benthic offshore biodiversity 3 1.3 Objective 2: Marine mammal research 4 1.4 Objective 3: Coastal biodiversity and connectivity 5 2. METHODS 5 2.1 Survey area 5 2.2 Survey design 6 Offshore Biodiversity 6 Marine mammal sampling 8 Coastal survey 8 Station recording 8 2.3 Sampling operations 8 Multibeam mapping 8 Photographic transect survey 9 Fish and Invertebrate sampling 9 Plankton sampling 11 Catch processing 11 Environmental sampling 12 Marine mammal sampling 12 Dive sampling operations 12 Outreach 13 3. -

Most Impaired" Coral Reef Areas in the State of Hawai'i
Final Report: EPA Grant CD97918401-0 P. L. Jokiel, K S. Rodgers and Eric K. Brown Page 1 Assessment, Mapping and Monitoring of Selected "Most Impaired" Coral Reef Areas in the State of Hawai'i. Paul L. Jokiel Ku'ulei Rodgers and Eric K. Brown Hawaii Coral Reef Assessment and Monitoring Program (CRAMP) Hawai‘i Institute of Marine Biology P.O.Box 1346 Kāne'ohe, HI 96744 Phone: 808 236 7440 e-mail: [email protected] Final Report: EPA Grant CD97918401-0 April 1, 2004. Final Report: EPA Grant CD97918401-0 P. L. Jokiel, K S. Rodgers and Eric K. Brown Page 2 Table of Contents 0.0 Overview of project in relation to main Hawaiian Islands ................................................3 0.1 Introduction...................................................................................................................3 0.2 Overview of coral reefs – Main Hawaiian Islands........................................................4 1.0 Ka¯ne‘ohe Bay .................................................................................................................12 1.1 South Ka¯ne‘ohe Bay Segment ...................................................................................62 1.2 Central Ka¯ne‘ohe Bay Segment..................................................................................86 1.3 North Ka¯ne‘ohe Bay Segment ....................................................................................94 2.0 South Moloka‘i ................................................................................................................96 2.1 Kamalō -

Langston R and H Spalding. 2017
A survey of fishes associated with Hawaiian deep-water Halimeda kanaloana (Bryopsidales: Halimedaceae) and Avrainvillea sp. (Bryopsidales: Udoteaceae) meadows Ross C. Langston1 and Heather L. Spalding2 1 Department of Natural Sciences, University of Hawai`i- Windward Community College, Kane`ohe,¯ HI, USA 2 Department of Botany, University of Hawai`i at Manoa,¯ Honolulu, HI, USA ABSTRACT The invasive macroalgal species Avrainvillea sp. and native species Halimeda kanaloana form expansive meadows that extend to depths of 80 m or more in the waters off of O`ahu and Maui, respectively. Despite their wide depth distribution, comparatively little is known about the biota associated with these macroalgal species. Our primary goals were to provide baseline information on the fish fauna associated with these deep-water macroalgal meadows and to compare the abundance and diversity of fishes between the meadow interior and sandy perimeters. Because both species form structurally complex three-dimensional canopies, we hypothesized that they would support a greater abundance and diversity of fishes when compared to surrounding sandy areas. We surveyed the fish fauna associated with these meadows using visual surveys and collections made with clove-oil anesthetic. Using these techniques, we recorded a total of 49 species from 25 families for H. kanaloana meadows and surrounding sandy areas, and 28 species from 19 families for Avrainvillea sp. habitats. Percent endemism was 28.6% and 10.7%, respectively. Wrasses (Family Labridae) were the most speciose taxon in both habitats (11 and six species, respectively), followed by gobies for H. kanaloana (six Submitted 18 November 2016 species). The wrasse Oxycheilinus bimaculatus and cardinalfish Apogonichthys perdix Accepted 13 April 2017 were the most frequently-occurring species within the H. -

Reef Fishes of the Bird's Head Peninsula, West
Check List 5(3): 587–628, 2009. ISSN: 1809-127X LISTS OF SPECIES Reef fishes of the Bird’s Head Peninsula, West Papua, Indonesia Gerald R. Allen 1 Mark V. Erdmann 2 1 Department of Aquatic Zoology, Western Australian Museum. Locked Bag 49, Welshpool DC, Perth, Western Australia 6986. E-mail: [email protected] 2 Conservation International Indonesia Marine Program. Jl. Dr. Muwardi No. 17, Renon, Denpasar 80235 Indonesia. Abstract A checklist of shallow (to 60 m depth) reef fishes is provided for the Bird’s Head Peninsula region of West Papua, Indonesia. The area, which occupies the extreme western end of New Guinea, contains the world’s most diverse assemblage of coral reef fishes. The current checklist, which includes both historical records and recent survey results, includes 1,511 species in 451 genera and 111 families. Respective species totals for the three main coral reef areas – Raja Ampat Islands, Fakfak-Kaimana coast, and Cenderawasih Bay – are 1320, 995, and 877. In addition to its extraordinary species diversity, the region exhibits a remarkable level of endemism considering its relatively small area. A total of 26 species in 14 families are currently considered to be confined to the region. Introduction and finally a complex geologic past highlighted The region consisting of eastern Indonesia, East by shifting island arcs, oceanic plate collisions, Timor, Sabah, Philippines, Papua New Guinea, and widely fluctuating sea levels (Polhemus and the Solomon Islands is the global centre of 2007). reef fish diversity (Allen 2008). Approximately 2,460 species or 60 percent of the entire reef fish The Bird’s Head Peninsula and surrounding fauna of the Indo-West Pacific inhabits this waters has attracted the attention of naturalists and region, which is commonly referred to as the scientists ever since it was first visited by Coral Triangle (CT). -

My Friend, the Volcano (Adapted from the 2004 Submarine Ring of Fire Expedition)
New Zealand American Submarine Ring of Fire 2007 My Friend, The Volcano (adapted from the 2004 Submarine Ring of Fire Expedition) FOCUS MAXIMUM NUMBER OF STUDENTS Ecological impacts of volcanism in the Mariana 30 and Kermadec Islands KEY WORDS GRADE LEVEL Ring of Fire 5-6 (Life Science/Earth Science) Asthenosphere Lithosphere FOCUS QUESTION Magma What are the ecological impacts of volcanic erup- Fault tions on tropical island arcs? Transform boundary Convergent boundary LEARNING OBJECTIVES Divergent boundary Students will be able to describe at least three Subduction beneficial impacts of volcanic activity on marine Tectonic plate ecosystems. BACKGROUND INFORMATION Students will be able to explain the overall tec- The Submarine Ring of Fire is an arc of active vol- tonic processes that cause volcanic activity along canoes that partially encircles the Pacific Ocean the Mariana Arc and Kermadec Arc. Basin, including the Kermadec and Mariana Islands in the western Pacific, the Aleutian Islands MATERIALS between the Pacific and Bering Sea, the Cascade Copies of “Marianas eruption killed Anatahan’s Mountains in western North America, and numer- corals,” one copy per student or student group ous volcanoes on the western coasts of Central (from http://www.cdnn.info/eco/e030920/e030920.html) America and South America. These volcanoes result from the motion of large pieces of the AUDIO/VISUAL MATERIALS Earth’s crust known as tectonic plates. None Tectonic plates are portions of the Earth’s outer TEACHING TIME crust (the lithosphere) about 5 km thick, as Two or three 45-minute class periods, plus time well as the upper 60 - 75 km of the underlying for student research mantle. -

Patterns of Prehistoric Human Mobility in Polynesia Indicated by Mtdna from the Pacific Rat (Rattus Exulans͞population Mobility)
Proc. Natl. Acad. Sci. USA Vol. 95, pp. 15145–15150, December 1998 Anthropology Patterns of prehistoric human mobility in Polynesia indicated by mtDNA from the Pacific rat (Rattus exulansypopulation mobility) E. MATISOO-SMITH*†,R.M.ROBERTS‡,G.J.IRWIN*, J. S. ALLEN*, D. PENNY§, AND D. M. LAMBERT¶ *Department of Anthropology and ‡School of Biological Sciences, University of Auckland, P. B. 92019 Auckland, New Zealand; and §Molecular Genetics Unit and ¶Department of Ecology, Massey University, P. B. 11222 Palmerston North, New Zealand Communicated by R. C. Green, University of Auckland, Auckland, New Zealand, October 14, 1998 (received for review July 20, 1998) ABSTRACT Human settlement of Polynesia was a major Recent genetic research focusing on Polynesian populations event in world prehistory. Despite the vastness of the distances has contributed significantly to our understanding of the covered, research suggests that prehistoric Polynesian popu- ultimate origins of this last major human migration. Studies of lations maintained spheres of continuing interaction for at globin gene variation (2) and mtDNA lineages of modern least some period of time in some regions. A low level of genetic Polynesians (3, 4) and studies of ancient DNA from Lapita- variation in ancestral Polynesian populations, genetic admix- associated skeletons (5) may indicate that some degree of ture (both prehistoric and post-European contact), and severe admixture with populations in Near Oceania occurred as more population crashes resulting from introduction of European remote biological ancestors left Southeast Asia and passed diseases make it difficult to trace prehistoric human mobility through Near Oceania. An alternative hypothesis is that the in the region by using only human genetic and morphological biological ancestors of these groups were one of a number of markers. -

Venom Evolution Widespread in Fishes: a Phylogenetic Road Map for the Bioprospecting of Piscine Venoms
Journal of Heredity 2006:97(3):206–217 ª The American Genetic Association. 2006. All rights reserved. doi:10.1093/jhered/esj034 For permissions, please email: [email protected]. Advance Access publication June 1, 2006 Venom Evolution Widespread in Fishes: A Phylogenetic Road Map for the Bioprospecting of Piscine Venoms WILLIAM LEO SMITH AND WARD C. WHEELER From the Department of Ecology, Evolution, and Environmental Biology, Columbia University, 1200 Amsterdam Avenue, New York, NY 10027 (Leo Smith); Division of Vertebrate Zoology (Ichthyology), American Museum of Natural History, Central Park West at 79th Street, New York, NY 10024-5192 (Leo Smith); and Division of Invertebrate Zoology, American Museum of Natural History, Central Park West at 79th Street, New York, NY 10024-5192 (Wheeler). Address correspondence to W. L. Smith at the address above, or e-mail: [email protected]. Abstract Knowledge of evolutionary relationships or phylogeny allows for effective predictions about the unstudied characteristics of species. These include the presence and biological activity of an organism’s venoms. To date, most venom bioprospecting has focused on snakes, resulting in six stroke and cancer treatment drugs that are nearing U.S. Food and Drug Administration review. Fishes, however, with thousands of venoms, represent an untapped resource of natural products. The first step in- volved in the efficient bioprospecting of these compounds is a phylogeny of venomous fishes. Here, we show the results of such an analysis and provide the first explicit suborder-level phylogeny for spiny-rayed fishes. The results, based on ;1.1 million aligned base pairs, suggest that, in contrast to previous estimates of 200 venomous fishes, .1,200 fishes in 12 clades should be presumed venomous. -

Obsidian from Macauley Island: a New Zealand Connection
www.aucklandmuseum.com Obsidian from Macauley Island: a New Zealand connection Louise Furey Auckland War Memorial Museum Callan Ross-Sheppard McGill University, Montreal Kath E. Prickett Auckland War Memorial Museum Abstract An obsidian flake collected from Macauley Island during the Kermadec Expedition has been analysed to determine the source. The result indicates a Mayor Island, New Zealand, origin, supporting previous results on obsidian from Raoul Island that Polynesians travelled back into the Pacific from New Zealand. Keywords obsidian; Pacific colonisation; Pacific voyaging; Macauley Island. INTRODUCTION The steep terrain meant there were only a few suitable habitation places confined to flat areas of limited size on The Kermadec Islands comprise Raoul Island, the largest the south and north coasts. The only landing places were of the group, the nearby Meyer Islands and Herald adjacent to these flats. Botanical surveys have recorded Islets, and 100–155 km to the south are Macauley, the presence of Polynesian tropical cultigens including Curtis and Cheeseman iIslands, and L’Esperence Rock. candlenut (Aleurites monuccana) and ti (Cordyline Raoul is approximately 1000 km to the northeast of fruticosa) (Sykes 1977) which are not endemic to the New Zealand. During the 2011 Kermadec Expedition, island and it is assumed they were transported there by samples of naturally occurring obsidian were obtained Polynesian settlers. Polynesian tuberous vegetables such from Raoul, the Meyer Islands, and from the sea floor off as taro (Colocasia esculenta) and kumara (Ipomoea Curtis Island. These are now in the Auckland Museum batatas) were also recorded but their introduction collection. More importantly however was the single attributed to later 19th century immigrants. -

The Marine Biodiversity and Fisheries Catches of the Pitcairn Island Group
The Marine Biodiversity and Fisheries Catches of the Pitcairn Island Group THE MARINE BIODIVERSITY AND FISHERIES CATCHES OF THE PITCAIRN ISLAND GROUP M.L.D. Palomares, D. Chaitanya, S. Harper, D. Zeller and D. Pauly A report prepared for the Global Ocean Legacy project of the Pew Environment Group by the Sea Around Us Project Fisheries Centre The University of British Columbia 2202 Main Mall Vancouver, BC, Canada, V6T 1Z4 TABLE OF CONTENTS FOREWORD ................................................................................................................................................. 2 Daniel Pauly RECONSTRUCTION OF TOTAL MARINE FISHERIES CATCHES FOR THE PITCAIRN ISLANDS (1950-2009) ...................................................................................... 3 Devraj Chaitanya, Sarah Harper and Dirk Zeller DOCUMENTING THE MARINE BIODIVERSITY OF THE PITCAIRN ISLANDS THROUGH FISHBASE AND SEALIFEBASE ..................................................................................... 10 Maria Lourdes D. Palomares, Patricia M. Sorongon, Marianne Pan, Jennifer C. Espedido, Lealde U. Pacres, Arlene Chon and Ace Amarga APPENDICES ............................................................................................................................................... 23 APPENDIX 1: FAO AND RECONSTRUCTED CATCH DATA ......................................................................................... 23 APPENDIX 2: TOTAL RECONSTRUCTED CATCH BY MAJOR TAXA ............................................................................ -
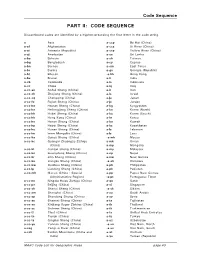
Code Sequence
Code Sequence PART II: CODE SEQUENCE Discontinued codes are identified by a hyphen preceding the first letter in the code string. a Asia a-ccp Bo Hai (China) a-af Afghanistan a-ccs Xi River (China) a-ai Armenia (Republic) a-ccy Yellow River (China) a-aj Azerbaijan a-ce Sri Lanka a-ba Bahrain a-ch Taiwan a-bg Bangladesh a-cy Cyprus a-bn Borneo a-em East Timor a-br Burma a-gs Georgia (Republic) a-bt Bhutan -a-hk Hong Kong a-bx Brunei a-ii India a-cb Cambodia a-io Indonesia a-cc China a-iq Iraq a-cc-an Anhui Sheng (China) a-ir Iran a-cc-ch Zhejiang Sheng (China) a-is Israel a-cc-cq Chongqing (China) a-ja Japan a-cc-fu Fujian Sheng (China) a-jo Jordan a-cc-ha Hainan Sheng (China) a-kg Kyrgyzstan a-cc-he Heilongjiang Sheng (China) a-kn Korea (North) a-cc-hh Hubei Sheng (China) a-ko Korea (South) a-cc-hk Hong Kong (China) a-kr Korea a-cc-ho Henan Sheng (China) a-ku Kuwait a-cc-hp Hebei Sheng (China) a-kz Kazakhstan a-cc-hu Hunan Sheng (China) a-le Lebanon a-cc-im Inner Mongolia (China) a-ls Laos a-cc-ka Gansu Sheng (China) -a-mh Macao a-cc-kc Guangxi Zhuangzu Zizhiqu a-mk Oman (China) a-mp Mongolia a-cc-ki Jiangxi Sheng (China) a-my Malaysia a-cc-kn Guangdong Sheng (China) a-np Nepal a-cc-kr Jilin Sheng (China) a-nw New Guinea a-cc-ku Jiangsu Sheng (China) -a-ok Okinawa a-cc-kw Guizhou Sheng (China) a-ph Philippines a-cc-lp Liaoning Sheng (China) a-pk Pakistan a-cc-mh Macao (China : Special a-pp Papua New Guinea Administrative Region) -a-pt Portuguese Timor a-cc-nn Ningxia Huizu Zizhiqu (China) a-qa Qatar a-cc-pe Beijing (China) a-si Singapore -

Conservation Status of New Zealand Marine Mammals, 2019 Report
2019 NEW ZEALAND THREAT CLASSIFICATION SERIES 29 Conservation status of New Zealand marine mammals, 2019 C.S. Baker, L. Boren, S. Childerhouse, R. Constantine, A. van Helden, D. Lundquist, W. Rayment and J.R. Rolfe Dedication This report is dedicated to the memory of Dr Rob Mattlin, a founding member of this Committee, who passed away in May 2019. Rob’s knowledge, insight and personality will be greatly missed by the Committee and by the marine mammals for which he was so passionate. Rob made New Zealand his home after leaving his native USA to undertake a PhD on New Zealand fur seals at Canterbury University. He proceeded to dedicate his life to marine science with a particular focus on marine mammals and spent most of his time based in New Zealand, apart from a role at the US Marine Mammal Commission. His contribution to our understanding of marine mammal biology and ecology and, in particular, his expertise in interactions with fisheries, was a key factor in achieving many positive outcomes for marine mammals in New Zealand. He made significant contributions to the conservation and management of both New Zealand sea lions and Hector’s dolphins. The Committee would like to recognise the unique balance of wisdom, humour, and tenacity that Rob brought to his work and hope that we can continue to match his enthusiasm and passion. Cover: The southern right whale (Eubalaena australis) population is recovering from perilously low numbers and is now assessed as At Risk – Recovering, an improvement on its previous Threatened status. Photo: Will Rayment. -

61661147.Pdf
Resource Inventory of Marine and Estuarine Fishes of the West Coast and Alaska: A Checklist of North Pacific and Arctic Ocean Species from Baja California to the Alaska–Yukon Border OCS Study MMS 2005-030 and USGS/NBII 2005-001 Project Cooperation This research addressed an information need identified Milton S. Love by the USGS Western Fisheries Research Center and the Marine Science Institute University of California, Santa Barbara to the Department University of California of the Interior’s Minerals Management Service, Pacific Santa Barbara, CA 93106 OCS Region, Camarillo, California. The resource inventory [email protected] information was further supported by the USGS’s National www.id.ucsb.edu/lovelab Biological Information Infrastructure as part of its ongoing aquatic GAP project in Puget Sound, Washington. Catherine W. Mecklenburg T. Anthony Mecklenburg Report Availability Pt. Stephens Research Available for viewing and in PDF at: P. O. Box 210307 http://wfrc.usgs.gov Auke Bay, AK 99821 http://far.nbii.gov [email protected] http://www.id.ucsb.edu/lovelab Lyman K. Thorsteinson Printed copies available from: Western Fisheries Research Center Milton Love U. S. Geological Survey Marine Science Institute 6505 NE 65th St. University of California, Santa Barbara Seattle, WA 98115 Santa Barbara, CA 93106 [email protected] (805) 893-2935 June 2005 Lyman Thorsteinson Western Fisheries Research Center Much of the research was performed under a coopera- U. S. Geological Survey tive agreement between the USGS’s Western Fisheries